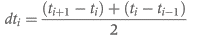Резюме
Шиншилла (Chinchilla laniger) - растительноядный гистрикоморфный южноамериканский грызун, для которого до настоящего времени не сообщалось о среднем временем удержания содержимого желудка. Шести животным (стандартное отклонение средней массы тела: 513 ± 99 г) на диете из травяного сена и гранул на основе люцерны давали дозу растворенного вещества (кобальт-ЭДТА) и частиц (волокно хромовой протравы, <2 мм) маркера с последующим частым сбором фекалий. Потребление сухого вещества составляло 45,2 ± 8,0 г / кг, 0,75 / день. Среднее время удерживания составляло 22,2 ± 5,3 ч для растворенных веществ и 25,4 ± 5,2 ч для частиц, причем разница между особями не была значимой. Это указывает на наличие механизма сепарции толстого кишечника по типу «ловушки слизью», который соответствует морфологическим описаниям типичной борозды толстой кишки у шиншилл. В соответствии со стратегией разделения содержимого желудка толстым кишечником и образования цекотрофов, пики вторичной экскреции маркера указывали на случаи копрофагии, которые были разнесены приблизительно на 12 часов. Принимая во внимание, что эти времена удержания кажутся более длительными, чем показатели, о которых сообщалось для кроликов (Oryctolagus cuniculus) или морских свинок (Cavia procllus), было бы интересно сравнить пищеварительную эффективность шиншилл при высоких уровнях пищевых волокон с другими видами.
Ключевые слова: шиншилла, пищеварение, копрофагия, цекотрофия, травоядные, ферментация
Получено: 2 июня 2015; принято: 31 октября 2015
Введение
Копрофагия представляет собой пищеварительную стратегию, применяемую мелкими травоядными для уменьшения потенциальной потери питательных веществ, связанных с выделением микробного вещества, которое происходит в результате заднекишечной ферментации, с фекалиями (Карасов и Март Инез дель Рио, 2007). Для того, чтобы эта стратегия была эффективной, вещество, выделяемое в виде фекалий, должно быть разделено на часть, предназначенную для повторного потребления, и часть, которая действительно выводится. Механизм, обеспечивающий такое разделение, был назван «механизм разделения толстого кишечника» (CSM, Björnhag, 1972). Были задокументированы два принципиально разных таких механизма (Hume and Sakaguchi, 1991; Björnhag and Snipes, 1999; Franz et al., 2011). Тот, который присутствует у лагоморфов (кроликов и зайцев), связан с ретроградным вымыванием содержимого желудка в толстом кишечнике, которое «выталкивает» бактерии обратно в слепую кишку и называется «разделением вымывания». Тот, который присутствует у грызунов и называется «ловушкой слизью», основан на экстракции микробов из содержимого желудка в слизь, содержащуюся в особой структуре толстого кишечника, с которой захваченные микробы транспортируются обратно в слепую кишку. У гистрикоморфных («дикобразоподобных», включая морских свинок Cavia porcellus или шиншилл), грызунов эта специфическая структура представляет собой так называемую «борозду» толстого кишечника (Gorgas, 1966; Snipes et al., 1988) и широко изучалась (Takahashi и Sakaguchi, 2000, 2006). У миоморфных («мышевидных») грызунов этот механизм может быть связан с определенными продольными складками и косыми бороздами (Plicae circlares) в толстом кишечнике (Behmann, 1973; Sperber et al., 1983), но детально не исследован. Из слепой кишки материал, отделенный CSM, выводится в виде цекотрофов, переваривается животным и подвергается ферментативному пищеварению в желудке и тонком кишечнике. Соответствующий механизм у землероек, которые также практикуют копрофагию (Loxton et al., 1975), не был идентифицирован.
Эксперименты по прохождению содержимого желудка, в которых неперевариваемый маркер проглатывается животными в виде импульсной дозы, а его концентрация впоследствии регистрируется в фекалиях с течением времени, могут продемонстрировать само по себе копрофагальное поведение из-за вторичных пиков маркера, указывающих на повторное поглощение маркера с фекалиями (Clauss et al. al., 2007). Вторичные пики не обязательно возникают, так как повторное потребление микробных компонентов может не потребоваться, если сама диета предлагает питательные вещества, в частности белок, в высоких концентрациях. Таким образом, было показано, что копрофагия варьируется в зависимости от рациона у кроликов (Oryctolagus cuniculus) (Fekete и Bokori, 1985; Garcıa et al., 1995) или вискач (Lagostomus maximus) (Hagen et al., 2015a). Эксперименты с прохождением могут дополнительно дифференцировать CSM «вымывания», при котором маркеры растворенных веществ в частности повторно используются посредством копрофагии и, следовательно, имеют намного более длительное время удержания, чем маркеры частиц, и CSM «ловушка слизью», в котором маркеры растворенных веществ и частицы перемещаются вместе (Hume и Sakaguchi, 1991; Pei et al., 2001; Franz et al., 2011).
Шиншиллы (Chinchilla laniger) - травоядные южноамериканские грызуны. Их естественная диета в основном состоит из трав, а также растений, коры кустарников, бромелий или плодов кактусов, в зависимости от наличия соответствующих растений (Cortes et al., 2002). Шиншиллы имеют долгую историю содержания как пушные звери, так и домашние животные. Рекомендации по кормлению включают предоставление источника грубого корма, гранул с высоким содержанием клетчатки и лишь ограниченного количества свежей растительности или овощей (Wolf et al., 2003; Grant, 2014; Kohles, 2014). У шиншилл есть борозда толстого кишечника, типичная для гистрикоморфных грызунов (Gorgas, 1966), которая является неотъемлемой частью их CSM-«ловушки слизью» (Holtenius and Björnhag, 1985). Не указывая источник или метод, Johnson-Delaney (2006) утверждает, что время прохождения содержимого через желудочно-кишечный тракт шиншилл составляет 12 - 15 часов. Оценка среднего времени удержания содержимого в пищеварительной системе была оценена как 35 ч в исследовании с полным изъятием пищи, непрерывным нанесением маркера в течение 24 ч и длительными интервалами отбора проб кала (Krishnamurti et al., 1974), но пока что характерные особенности экскреции и среднее значение время удержания маркеров растворенных веществ и частиц у этого вида не исследовалось. Такие данные представляют особый интерес, потому что по сравнению с другими гистрикоморфными грызунами у шиншилл чрезвычайно длинный толстый кишечник (Gorgas, 1966). Это может быть адаптацией к засушливым местам обитания и, возможно, позволяет им жить при более низких уровнях потребления воды, чем другим грызунам (Hagen et al., 2014). Мы выдвинули гипотезу, что типы экскреции маркеров при прохождении будут указывать на копрофагию по вторичным пикам, CSM «ловушка слизью» при параллельном перемещении маркеров растворенного вещества и частиц и давать продолжительные значения среднего времени прохождения из-за длинного толстого кишечника у этого вида.
Материалы и методы
Шесть шиншилл (Chinchilla laniger; стандартное отклонение средней массы тела: 513 ± 99 г) использовались ранее в исследовании потребления пищи и воды (Hagen et al., 2014) (лицензия на эксперимент 80/2012). Животных содержали по отдельности в открытых ящиках с древесной стружкой в комнате с температурой от 22 до 26 ° С. Шиншилл кормили травяным сеном (г / кг сухого вещества: сырой белок, 221; волокно нейтрального детергента (клетчатка NDF), 588) и коммерческими гранулами на основе люцерны (г / кг сухого вещества: сырой белок, 139; NDF, 369) ad libitum. Потребление пищи определяли количественно, взвешивая количество предлагаемых продуктов питания и взвешивая остатки при следующем кормлении. Было измерено среднее время удержания маркера растворенного вещества (кобальт-ЭДТА) и маркера частиц (частицы волокна с хромовым покрытием <2 мм), полученные в соответствии с Uden et al. (1980). Концентрация хрома в протравленных волокнах составляла 37 г / кг. Маркеры скармливали в количестве 0,01 г Co-EDTA и 0,04 г протравленного волокна на животное в виде импульсной дозы в 18:00 ч, его смешивали с диетической формулой на основе сена (Critical CareOxbow Animal Health, Murdock NE) и вводили перорально шприцом животным, удерживаемым вручную (вынимались из вольера). Была предпринята осторожность, чтобы стереть всю непоглощенную смесь маркеров с морд животных и избежать переноса смеси маркеров в вольер при помещении животных обратно. Отбор проб фекалий проводился каждые 4 часа в течение первых 48 часов, каждые 6 часов, следующие 48 часов и один раз в пятый день. Фекалии анализировали на маркеры после влажного озоления с помощью атомно-абсорбционной спектроскопии согласно Behrend et al. (2004). Образцы, которые отклонялись от общей картины снижения концентрации маркера после начального пика (то есть образцы, позднее интерпретированные как «вторичные пики»), анализировали в двух экземплярах, если были доступны достаточные количества. Среднее время прохождения через весь пищеварительный тракт рассчитывали в соответствии с Thielemans et al. (1978) как
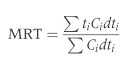
Где Ci = концентрация маркера в образцах фекалий из интервала времени ti (часы после введения маркера, с использованием средней точки интервала выборки), и dt i = интервал (в часах) соответствующей выборки.
Предполагалось, что маркер полностью выводится из организма после того, как концентрации его в фекалиях были сходны с фоновыми уровнями, определенными в образцах фекалий до введения дозы. Среднее время прохождения двух маркеров обычно распределяли и сравнивали с помощью парного t-критерия с использованием SPSS 21.0 (SPSS, Chicago, IL, USA).
Результаты
Относительное среднее потребление сухого вещества (на единицу метаболической массы тела) составляло 45,2 ± 8,1 (диапазон 34–55) г / кг 0,75 / день. Время прохождения (время появления первого маркера в фекалиях) составило всего 4 ч у некоторых особей и 8 или 12 ч у других (рис. 1). Среднее значение времени прохождения для маркера растворенного вещества (Co) составляло 22,2 ± 5,3 (диапазон 16,3-30,2) ч, а для маркера частиц (Cr) 25,4 ± 5,2 (диапазон 19,5 - 34,1) ч. Тип экскреции характеризовался 2–4 пиками маркера (рис. 1), которые иногда располагались на расстоянии всего 12 ч друг от друга, в частности первый и второй пики. Два маркера двигались в основном параллельно. Разница между ними не была значимой на уровне 0,05, хотя тенденция к более длительному удержанию частиц была очевидной (р = 0,082). Скорее всего, это произошло из-за двух животных, у которых большая часть маркеров частиц выводилась только после первого акта копрофагии (рис. 1 c, f).
Обсуждение
Результаты подтверждают, что у шиншиллы есть «ловушка слизью» и практикуется копрофагия, поведение, которое трудно наблюдать непосредственно. Что касается точной величины времени удержания содержимого желудка у шиншилл, то необходимы дополнительные эксперименты с использованием различных диет и уровней потребления для подтверждения этого. Что касается используемых маркеров, следует проверить, действительно ли протравленные волокна являются репрезентативными для неперевариваемого содержимого ЖКТ у грызунов, даже если широкое использование этих маркеров облегчает включение результатов настоящего исследования в более крупные наборы данных (например, Muller et al., 2013). В частности, идентификация вторичных пиков маркеров может быть яблоком раздора, поскольку можно спорить, являются ли определенные пики просто отдельными артефактами или действительно отражают акты копрофагии. Следует проявлять осторожность, чтобы избежать загрязнения маркерами во время их введения, а для критических образцов следует выполнить повторный анализ, как тот, который выполнен в данном исследовании. Это может повысить надежность интерпретации только до определенной степени. В частности, что касается предполагаемых вторичных пиков с большим расстоянием до времени введения маркера, которые, например, отмечены на рис. 1a, e или f, было бы целесообразно поддерживать более частый и трудозатратный режим отбора проб кала, что обычно применяется только в течение первых двух дней в исследовании прохождения. События, которые отмечены не только одним пиковым значением, но и несколькими образцами, указывающими на повторяющееся увеличение и уменьшение концентрации маркера (как, например, видно из первых пиков на рис. 1e), могут более надежно интерпретироваться как отсутствие артефактов. Однако следует отметить, что повторяющиеся пики маркеров также были задокументированы у особей видов, у которых не известно о копрофагии, а загрязнение фекалий маркерным субстратом может быть исключено и повторное попадание фекалий во время эксперимента не наблюдалось (например, Matsuda et al., 2015). Следовательно, типы экскреции маркеров всегда следует рассматривать в контексте других морфологических и физиологических данных. Учитывая различия в том, были ли сходные или разнородные концентрации обоих маркеров выделены на вторичных пиках, следует сделать вывод, что механизм разделения в виде «ловушки слизью» не демонстрирует явного предпочтения ни для маркера растворенного вещества, ни для маркера частиц, и, возможно, степень вероятности связана со степенью сохранения маркера. Интервал между повторными пиками экскреции маркера предполагает ритм копрофагии, который короче, чем 24 часа, и ближе к 12 часам. Это совпадает с результатами Holtenius и Björnhag (1985), которые наблюдали отдельные случаи копрофагии у шиншилл в течение всего дня (но не ночью). Аналогичным образом, расстояние между пиками экскреции маркеров у вискач предполагает наличие актов копрофагии с 12-часовыми интервалами времени (Clauss et al., 2007). Кроме того, некоторые расстояния между пиками экскреции маркеров у кроликов и морских свинок были примерно одинаковыми (Sakaguchi et al., 1987; Franz et al., 2011). Однако у кроликов также отмечены пики маркеров, разнесенные на 24 часа (Sakaguchi et al., 1987). По-видимому, ритм актов копрофагии варьируется между видами. Некоторые грызуны практикуют копрофагию несколько раз в течение дня, например, травоядная полевка (Microtus californicus) (Kenagy and Hoyt, 1979), луговая полевка (M. pennsylvanicus) (Ouellette and Heisinger, 1980), норвежский лемминг (Lemmus lemmus) (Björnhag, 1977) или горный бобр (Aplodontia rufa) (Ingles, 1961). Дегу (Octodon degu) - другое животное, практикующее копрофагию примерно каждые 12 часов, но, в отличие от шиншиллы, дегу демонстрирует такое поведение только ночью (Kenagy et al., 1999). У зайцев-беляков (Lepus timidus) (Pehrson, 1983), кенгуровых крыс (Dipodomys microps) (Kenagy и Hoyt, 1979), нутрий (Myocastor coypus) (Gosling, 1979) и капибар (Hydrochoerus hydrochaeris) (Herrera, 1985) наблюдался ритм копрофагии 24 ч. Помимо этих очевидных видовых различий, вариации в составе питательных веществ и количестве доступной пищи могут изменить копрофагическое поведение внутри видов (Fekete и Bokori, 1985; Garcıa et al., 1995; Kenagy et al., 1999; Hagen et al. al., 2015a).
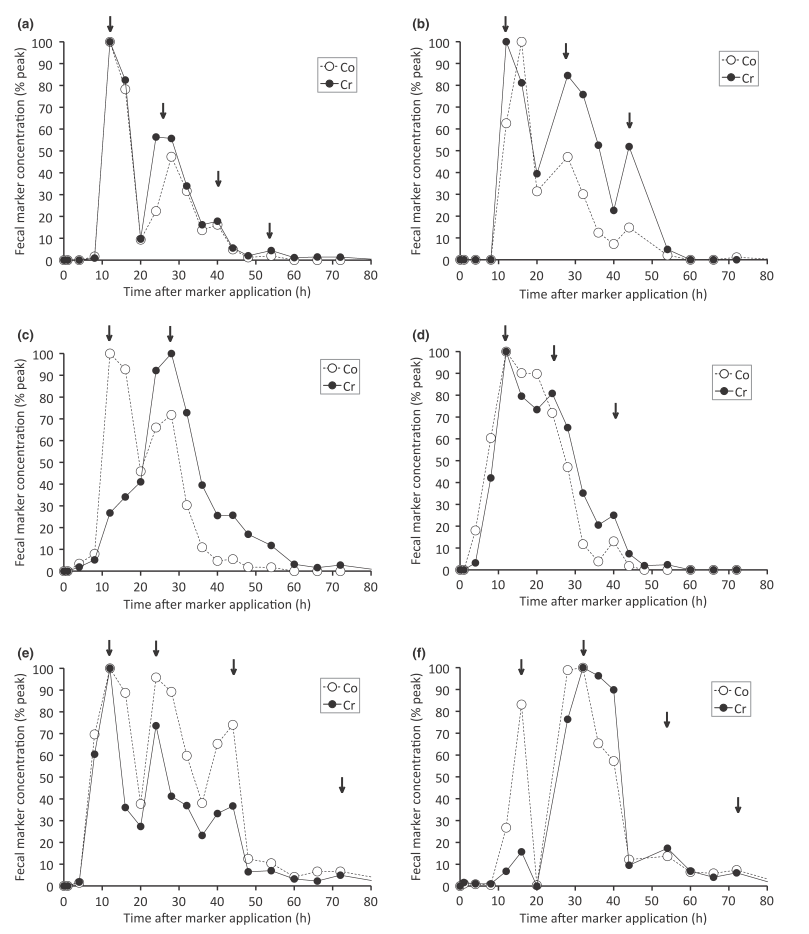
Рис. 1 Маркеры экскреции маркера растворенного вещества (кобальт-ЭДТА) и маркера частиц (волокно хромовой протравы, <2 мм) у шести шиншилл (Chinchilla laniger, a - e) в данном исследовании. Обратите внимание на повторяющиеся пики маркеров (отмечены стрелками), указывающие на копрофагию и общее отсутствие различий паттернов между двумя маркерами у отдельных особей.
Прим. пер. к рис. 1: Fecal marker concentration (% peak) – концентрация маркера в фекалиях (%, пик), Time after application (h) – время после введения маркера (ч), Co – кобальт, Cr – хром.
В отличие от среднего времени прохождения у кроликов (около 15 часов) и морских свинок (около 18 часов) с одинаковыми маркерами при питании, содержащем только травяное сено (Franz et al., 2011), у шиншилл наблюдается удивительно длинное время прохождения для их размера тела, достигающее в нижнем диапазоне значений лошадей, питавшихся только травяным сеном, с той же системой маркеров (23 - 34 ч, Clauss et al., 2014). Дает ли это более высокую усвояемость клетчатки у шиншилл, чем, например, у кроликов и морских свинок, или это просто побочный эффект того факта, что у шиншилл, как у гистрикоморфных грызунов, особенно длинный толстый кишечник для реабсорбции воды в качестве адаптации к засушливой среде обитания (Gorgas, 1966), ещё предстоит выяснить. Отдельные измерения переваримости, о которых сообщалось для шиншилл, не позволяют окончательно сказать, в равной степени или меньше влияет диета на эффективность переваривания клетчатки у шиншилл по сравнению с кроликами или морскими свинками (Hagen et al., 2015b). Сообщения о том, что домашние шиншиллы страдают от потери массы тела, когда их держат на диете с травяным сеном или только из свежей травы (Wolf et al., 2003), по-видимому, не соответствуют высокой усвояемости, которую можно интуитивно связать с длительным временем удерживания. Хотя копрофагия понимается главным образом как механизм рециркуляции питательных веществ, связанных с желудочно-кишечными бактериями (обозревается в Karasov and Martınez del Rio, 2007; Franz et al., 2011), тот факт, что маркированные волокна в настоящем исследовании и аналогичных исследованиях также рециркулируют по этому механизму, предполагает, что некоторые волокнистые компоненты также будут подвергаться вторичной и, следовательно, более длительной ферментации в желудочно-кишечном тракте при заднекишечной ферментации с помощью механизма разделения «ловушка слизью».
Благодарности
Мы благодарим профессора Макса М. Виттенбринка за использование оборудования, Зину Биггер за предоставление нам шиншилл, Хайдруна Барлебена и Кармен Кунц за анализ образцов и двух анонимных рецензентов за комментарии к рукописи. Это исследование было частью проекта 310030_135252 / 1, финансируемого Швейцарским национальным научным фондом.
Использованные источники
Behmann, H., 1973: Vergleichend- und funktionell-anatomische Untersuchungen am Caecum und Colon myomorpher Nagetiere. Zeitschrift f € ur Wissenschaftliche Zoologie 186 , 173 – 294.
Behrend, A.; Lechner-Doll, M.; Streich, W. J.; Clauss, M., 2004: Seasonal faecal excretion, gut fill, liquid and particle marker retention in mouflon (Ovis ammon musimon), and a comparison with roe deer (Capreolus capreolus). Acta Theriologica 49 , 503 – 515.
Bjornhag, G., 1972: Separation and delay of contents in the rabbit colon. Swedish Journal of Agricultural Research 2 , 125 – 136.
Bjornhag, G., 1977: Demonstration of coprophagy in some rodents. Swedish Journal of Agricultural Research 7 , 105 – 113.
Bjornhag, G.; Snipes, R. L., 1999: Colonic separation mechanism in lagomorph and rodent species - a comparison. Zoosystematics and Evolution 75 , 275 – 281.
Clauss, M.; Besselmann, D.; Schwarm, A.; Ortmann, S.; Hatt, J.-M., 2007: Demon- strating coprophagy with passage markers? The example of the plains viscacha (Lagostomus maximus). Comparative Biochemistry and Physiology A 147 , 453 – 459.
Clauss, M.; Schiele, K.; Ortmann, S.; Fritz, J.; Codron, D.; Hummel, J.; Kienzle, E., 2014: The effect of very low food intake on digestive physiology and forage digestibility in horses. Journal of Animal Physiology and Animal Nutrition 98 , 107 – 118.
Cortes, A.; Miranda, E.; Jimenez, J. E., 2002: Seasonal food habits of the endangered long-tailed chinchilla (Chinchilla lanigera): the effect of precipitation. Mammalian Biology 67,167 – 175.
Fekete, S.; Bokori, J., 1985: The effect of the fiber and protein level of the ration upon the cecotrophy of rabbit. Journal of Applied Rabbit Research 8 , 68 – 71.
Franz, R.; Kreuzer, M.; Hummel, J.; Hatt, J.-M.; Clauss, M., 2011: Intake, selection, digesta retention, digestion and gut fill of two coprophageous species, rab- bits (Oryctolagus cuniculus) and guinea pigs (Cavia porcellus), on a hay-only diet. Journal of Animal Physiology and Animal Nutrition 95 , 564 – 570.
Garcıa, J.; De Blas, J. C.; Carabano, R.;Garcia, P., 1995: Effect of type of lucerne hay on caecal fermentation and nitrogen contribution through caecotrophy in rabbits. Reproduction Nutrition Development 35 , 267 – 275.
Gorgas, M., 1966: Vergleichendanatomische Untersuchungen am Magen-Darm-Kanal der Sciuromorpha, Hystricomorpha und Caviomorpha(Rodentia). Zeitschrift fuer Wissenschaftliche Zoologie 175 , 237 – 404.
Gosling, L. M., 1979: The twenty-four hour activity cycle of captive coypus (Myocastor coypus). Journal of Zoology 187 , 341 – 367.
Grant, K., 2014: Rodent nutrition: digestive comparisons of 4 common rodent species. Veterinary Clinics of North America: Exotic Animal Practice 17 , 471 – 483.
Hagen, K.; Clauss, M.; Hatt, J.-M., 2014: Drinking preferences in chinchillas (Chinchilla laniger), degus (Octodon degu) and Guinea pigs (Cavia porcellus). Journal of Animal Physiology and Animal Nutrition 98 , 942 – 947.
Hagen, K. B.; Besselmann, D.; Cyrus-Eulenberger, U.; Vendl, C.; Ortmann, S.; Zingg, R.; Kienzle, E.; Kreuzer, M.; Hatt, J.-M.; Clauss, M., 2015a: Digestive physiology of the plains viscacha (Lagostomus maximus), a large herbivorous hystricomorph rodent. Zoo Biology 34 , 345 – 359.
Hagen, K. B.; Tschudin, A.; Liesegang, A.; Hatt, J.-M.; Clauss, M., 2015b: Organic matter and macromineral digestibility in domestic rabbits (Oryctolagus cuniculus) as compared to other hindgut fermenters. Journal of Animal Physiology and Animal Nutrition 99 , 1197 – 1209.
Herrera, E. A., 1985: Coprophagy in the capybara, Hydrochoerus hydrochoeris. Journal of Zoology 207 , 616 – 619.
Holtenius, K.; Bjornhag, G., 1985: The colonic separation mechanism in the guinea pig (Cavia porcellus) and the chinchilla (Chinchilla laniger). Comparative Biochemistry and Physiology A 82 , 537 – 542.
Hume, I. D.; Sakaguchi, E., 1991: Patterns of digesta flow and digestion in foregut and hindgut fermenters. In: T. Tsuda, Y. Saaski, R. Kawashima (eds), Physiological Aspects of Digestion and Metabolism in Ruminants. Academic Press, San Diego, pp. 427 – 451. Ingles, L. G., 1961: Reingestion in the mountain beaver. Journal of Mammalogy 42 , 411 – 412.
Johnson-Delaney, C. A., 2006: Anatomy and physiology of the rabbit and rodent gastrointestinal system. Proceedings of the Association of Avian Veterinarians 2006, 9 – 17.
Karasov, W. H.; Martınez del Rio, C., 2007: Physiological Ecology: How Animals Process Energy, Nutrients, and Toxins. Princeton University Press, Princeton, New Jersey, USA.
Kenagy, G. J.; Hoyt, D., 1979: Reingestion of feces in rodents and its daily rhythmicity. Oecologia 44 , 403 – 409.
Kenagy, G. J.; Veloso, C.; Bozinovic, F.,1999: Daily rhythms of food intake and feces reingestion in the degu, an herbivorous Chilean rodent: optimizing digestion through coprophagy. Physiological and Biochemical Zoology 72 , 78 – 86.
Kohles, M., 2014: Gastrointestinal anatomy and physiology of select exotic companion mammals. Veterinary Clinics of North America: Exotic Animal Practice 17 , 165 – 178.
Krishnamurti, C. R.; Kitts, W. D.; Smith, D. C., 1974: The digestion of carbohydrates in the chinchilla (Chinchilla lanigera). Canadian Journal of Zoology 52 , 1227 – 1233.
Loxton, R. G.; Raffaelli, D.; Begon, M., 1975: Coprophagy and the diurnal cycle of the common shrew, Sorex araneus. Journal of Zoology 177 , 449 – 453.
Matsuda, I.; Sha, J. C. M.; Ortmann, S.; Schwarm, A.; Grandl, F.; Caton, J.; Jens, W.; Kreuzer, M.; Marlena, D.; Hagen, K. B.; Clauss, M., 2015: Excretion patterns of solute and different-sized particle passage markers in foregut-fermenting proboscis monkey (Nasalis larvatus) do not indicate an adaptation for rumination. Physiology and Behavior 149 , 45 – 52.
Muller, D. W. H.; Codron, D.; Meloro, C.; Munn, A.; Schwarm, A.; Hummel, J.; Clauss, M., 2013: Assessing the Jarman-Bell Principle: scaling of intake, digestibility, retention time and gut fill with body mass in mammalian herbivores. Comparative Biochemistry and Physiology A 164 , 129 – 140.
Ouellette, D. E.; Heisinger, J. F., 1980: Reingestion of feces by Microtus pennsylvanicus. Journal of Mammalogy 61 , 366 – 368.
Pehrson, A., 1983: Caecotrophy in caged mountain hares (Lepus timidus). Journal of Zoology 199 , 563 – 574.
Pei, Y. X.; Wang, D. H.; Hume, I. D., 2001: Selective digesta retention and coprophagy in Brandt’s vole (Microtus brandti). Journal of Comparative Physiology B 171 , 457 – 464.
Sakaguchi, E.; Itoh, H.; Uchida, S.; Horigome, T., 1987: Comparison of fibre digestion and digesta retention time between rabbits, guinea-pigs, rats and hamsters. British Journal of Nutrition 58, 149 – 158.
Snipes, R. L.; Hornicke, H.; Bjornhag, G.; Stahl, W., 1988: Regional differences in hindgut structure and function in the nutria (Myocastor coypus). Cell Tissue Research 252 , 435 – 447.
Sperber, I.; Bjornhag, G.; Ridderstrale, Y., 1983: Function of proximal colon in lemming and rat. Swedish Journal of Agricultural Research 13 , 243 – 256.
Takahashi, T.; Sakaguchi, E., 2000: Role of the furrow of the proximal colon in the production of soft and hard feces in nutrias (Myocastor coypus). Journal of Comparative Physiology B 170 , 531 – 535.
Takahashi, T.; Sakaguchi, E., 2006: Transport of bacteria across and along the large intestinal lumen of guinea pigs. Journal of Comparative Physiology B 176 , 173 – 178.
Thielemans, M. F.; Francois, E.; Bodart, C.; Thewis, A., 1978: Mesure du transit gastrointestinal chez le porc a l’aide des radiolanthanides. Comparaison avec le mouton. Annales De Biologie Animale, Biochimie, Biophysique 18 , 237 – 247.
Ud é n, P.; Colucci, P. E.; Van Soest, P. J., 1980: Investigation of chromium, cerium and cobalt as markers in digesta. Rate of passage studies. Journal of the Science of Food and Agriculture 31 , 625 – 632.
Wolf, P.; Schroeder, A.; Wenger, A.; Kamphues, J., 2003: The nutrition of the chinchilla as a companion animal – basic data, influences and dependences. Journal of Animal Physiology and Animal Nutrition 87 , 129 – 133.
Авторы: K. B. Hagen1,*, M. T. Dittmann 1,2, S. Ortmann 3, M. Kreuzer2, J.-M. Hatt1and M. Clauss1
1 Clinic for Zoo Animals, Exotic Pets and Wildlife, Vetsuisse Faculty, University of Zurich, Zurich, Switzerland
2 Institute for Agricultural Sciences, ETH Zurich, Zurich, Switzerland and
3 Leibniz Institute for Zoo and Wildlife Research (IZW) Berlin, Berlin, Germany
Journal of Animal Physiology and Animal Nutrition
Перевод на русский язык: Д.В. Михайлов
Переводчик благодарит Е.К. Климочкину за помощь в редактировании
K. B. Hagen, M. T. Dittmann, S. Ortmann, M. Kreuzer, J.-M. Hatt and M. Clauss. Retention of solute and particle markers in the digestive tract of chinchillas (Chinchilla laniger) // J Anim Physiol Anim Nutr (Berl). 2016 Oct;100(5):801-6