Резюме. Соотношение 119 самцов на 100 живорожденных самок у шиншиллы значительно выше, чем у большинства других млекопитающих. Этот избыток рождающихся самцов может быть связан с необычайно выраженной разницей размеров X- и Y-хромосом.
Введение
Небольшое превышение числа рождений самцов у большинства млекопитающих отражает взаимодействие двух факторов: первичного соотношения полов при оплодотворении и возможного наличия половых различий при внутриутробной смертности. Одним из объяснений этого предполагаемого отклонения первичного соотношения полов от единицы является то, что сперматозоиды, несущие меньшую Y-хромосому, более подвижны, чем те, которые несут X-хромосому, и, следовательно, с большей вероятностью будут влиять на оплодотворение.
Шиншилла обладает X-хромосомами «дублированного типа», а её Y-хромосома является одной из самых маленьких в наборе (Galton, Benirschke & Ohno, 1965). Следовательно, разница в размерах между X- и Y-хромосомами гораздо больше, чем у большинства других млекопитающих. В связи с этим шиншилла является полезным видом для проверки того, становится ли разница размеров X- и Y-хромосом причиной избыточного рождения самцов. Если это предположение действительно верно, то преобладание потомства мужского пола у шиншиллы будет большим, чем у человека и других животных с более равными размерами X- и Y-хромосом.
Важный усложняющий фактор вносится в любое такое обсуждение, когда в одной колонии обнаруживается, что качество окружающего освещения оказало радикальное влияние на соотношение полов (Ott, 1964). При содержании в уличных вольерах при естественном освещении эти шиншиллы приносили равное количество самцов и самок. Но когда тех же родителей перевели в помещение и держали при обычном освещении лампами накаливания, рождались практически только самцы. Переход на дневной (синеватый) свет лампы накаливания привел к рождению практически одних только самок. Третий тип ламп накаливания («skyblue Coloramic» (один из типов ламп накаливания, производившихся в годы проведения исследования в США компанией General Electric – прим. пер.), излучающий как красный, так и синий свет, привел к рождению животных обоих полов.
В более раннем исследовании (Hillemann & Tibbitts, 1957), в котором, однако, не был указан тип освещения, соотношение полов 2586 новорожденных шиншилл составило 121,97 (самцов / 100 самок), что значительно выше, чем распространенное значение примерно 106 (самцов/100 самок) для многих других млекопитающих. В связи с этим было решено пересмотреть соотношение полов у шиншиллы.
Методы и результаты
Благодаря любезности редактора анкета была опубликована в торговом журнале заводчиков шиншилл (текст-рис. 1) (Galton, 1965). Семьдесят один заполненный вопросник был получен от владельцев шиншилловых ранчо, шестьдесят девять из США, Канады и Аляски, один из Англии и один из Южной Африки. Помимо данных о количестве живорожденных и мертворожденных, была также представлена информация о местонахождении колонии и типе освещения. Две анкеты были исключены из дальнейшего рассмотрения: в одной из них описывалось 52 рождения, но не указывалось количество детенышей каждого пола, а другая, в которой описывались 1286 рождений, из-за нескольких аномальных характеристик, в частности, очень низкого соотношения полов (36,5), необычно большого среднего размера помета (3,93) и удивительно низкого количества мертворожденных (1,6 мертворожденных на 1000 живорождений). В данных о соотношении полов, предоставленных в 69 анкетах, выбранных для исследования, отсутствовала значимая степень гетерогенности, согласно методу Брандта и Снедекора (Snedecor, 1956), (χ268DF= 83,96; 0,1>P> 0,05), в то время как включение данных из последней анкеты в анализ привело к высокозначимой степени гетерогенности (χ269DF = 433,85; P≤0,0005).

Рис.-текст 1. Вопросник соотношения полов шиншилл (Galton, 1965).
Ранчо находились в эксплуатации в течение периода от менее 1 года (девять ранчо) до 15 лет (три ранчо). Средняя продолжительность составила 4 года, 60% зарегистрированных ранчо работали 3 лет. Среднее количество живорождений на ранчо составило 155. Однако в половине заполненных анкет было зарегистрировано менее пятидесяти живорождений. Десять ранчо предоставили данные о 428-1065 живорожденных животных. В то время как большинство новых и мелких ранчо описывали свой опыт разведения в целом, более старые и более крупные ранчо часто приводили детали только своего последнего 1 или 2 лет работы. 10 664 зарегистрированных живорожденных были потомством 2984 самок в 5279 пометах; следовательно, среднее количество пометов на одну самку составляло 1,77, а средний размер помета – 2,02 живорожденных детеныша. Последнее значение хорошо согласуется с ранее сообщавшимся средним размером помета 1,88 живорожденных (Hillemann & Tibbitts, 1957). Количество пометов на одну самку ниже, чем значение 6,35, зарегистрированное ранее, по-видимому, из-за выборочности, вызванной как недавним созданием колоний, предоставляющих полные отчеты о разведении, так и исключением гораздо более ранних данных в случае более крупных ранчо.
Таблица 1. Влияние света на соотношение полов
|
Категория
|
Ранчо
|
Живорожденные
|
Соотношение полов (100 x M/F)
|
95% доверительный интервал*
|
|
Самцы (M)
|
Самки (F)
|
|
Общая группа
|
69
|
5798
|
4866
|
119,15
|
114,71-123,79
|
|
Не менее ¾ естественного освещения
|
46
|
4679
|
3936
|
118,88
|
113,95-124,03
|
|
Не менее ¾ искусственного освещение
|
13
|
516
|
454
|
113,66
|
100,22-129,02
|
|
Только искусственный свет: флуоресцентный, минимум ¾ (перекрывается с предыдущей группой)
|
6
|
366
|
304
|
120,39
|
103,49-140,37
|
|
Колония размещена в помещении
|
65
|
5077
|
4292
|
118,29
|
113,59-123,20
|
|
Колония размещалась как на улице, так
и в помещении
|
3
|
575
|
487
|
118,07
|
104,69-133,32
|
*Пределы доверительного интервала 95% были рассчитаны на основе среднеквадратической ошибки доли самцов. (p=M/(M+F)):
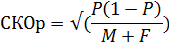
Значения P ± 1,96 СКОp выраженное как количество самцов / 100 самок:
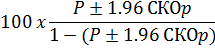
Соотношение полов для 10 664 живорожденных составило 119,15. Коэффициент мертворождаемости составил 76,95 на 1000 живорожденных. Соотношение полов у 302 мертворожденных, у которых был зарегистрирован пол, составило 134,11, что существенно не отличается от соотношения полов среди живорожденных (χ21DF = 0,892; 0,5>P> 0,3). Hillemann & Tibbitts (1957) сообщали о соотношении полов при рождении живых детенышей 121,97, о частоте 104,02 «дефектных» рождений на 1000 живорождений и о соотношении полов среди мертворождённых, значимо не отличавшемся от живорожденных. Значимые различия между соотношением полов в настоящем и прошлом исследовании отсутствуют, но разница между коэффициентом мертворождения в настоящей серии и частотой «дефектных» детенышей в более ранней работе весьма значима (P<0,001), возможно, из-за менее точного учета данных о мертворождениях в коммерческих колониях, предоставивших данные в нашей работе.
В таблице 1 показано соотношение полов в различных группах живорожденных в зависимости от местоположения колоний, находившихся в закрытых помещениях или на открытом воздухе, а также от типа освещения. Анкета была разработана для получения достаточно подробной информации о типе освещения. Однако 46 ранчо, на которые приходится 81% всех живорождений, использовали во время ухода за колониями в основном дневной свет, некоторые - исключительно, а некоторые - в сочетании электрическими лампами накаливания. Искусственный свет был преобладающим на тринадцати ранчо, семь из которых также использовали до четверти дневного света. Шесть ранчо, на которые приходится 6% всех живорожденных, имели только искусственное освещение, и в целом по крайней мере три четверти света обеспечивали обычные люминесцентные светильники, остальное - лампы накаливания. Ни одно из различий соотношения полов в любой из этих ситуаций не имеет значимых отличий ни от соотношения полов во всей группе, ни от значения в любой из других подгрупп. Точно так же не было значимых различий ни среди частот мертворождений, ни размеров помета, хотя их количество иногда было довольно мало.
Обсуждение
Соотношение полов среди 4098020 живорождений в США в 1963 г. составляло 105,27 (Национальный центр статистики здравоохранения США, 1966). Подобные значения были зарегистрированы у многих млекопитающих (Altman & Dittmer, 1962; Bacci, 1965). Соотношение полов у шиншиллы 119,15 значительно (P <0,001) превышает человеческое значение. Действительно, 95% доверительный интервал соотношения полов шиншиллы от 114,7 до 123,8 показывает, что шиншиллы обладают одним из самых высоких вторичных соотношений полов, поскольку имеется очень мало сообщений о соотношении полов такого порядка у других млекопитающих.
Ввиду отсутствия доказательств, относящихся к возможному существованию половых различий при внутриутробной смертности у шиншилл, разумно экстраполировать результаты недавних хромосомных исследований внутриутробного соотношения полов у других видов. У человека соотношение полов при 365 спонтанных и 404 индуцированных абортах, подвергнутых анализу кариотипа, составило 88,14 и 92,38 (Женевская конференция, 1966); Ни одно из этих значений значимо не отличается от соотношения полов при рождении у живорожденных людей в 1963 году, составлявшего 105,27. Предыдущие исследования внутриутробного соотношения полов у мужчин, основанные на морфологических критериях или критериях ядерно-полового хроматина, дали противоречивые результаты, предположительно отражающие неизбежные трудности, присущие этим методам (Crew, 1952; Stevenson, 1966). Однако тщательное морфологическое исследование самопроизвольных и искусственных абортов показало, что соотношение полов составляет 3003 мужчин : 2784 женщины, или 107,9 (Tietze, 1948). Хромосомный анализ восьмидесяти восьми 10-дневных бластоцист свиней показал соотношение полов среди животных, имевших нормальный кариотип, 38 XY : 39 XX (McFeely, 1967). Точно так же из девяноста восьми бластоцист мышей 49 имели мужскую конституцию половых хромосом, а 49 - женскую (Vickers, 1967). Из этих данных можно сделать вывод, что, по крайней мере, у человека, мыши и свиньи соотношение полов при рождении точно отражает соотношение полов при имплантации. Если предположить то же самое и для шиншиллы, это будет означать, что первичное соотношение полов у нее намного выше, чем у многих других млекопитающих.
К сожалению, невозможно исключить возможность систематической ошибки в данной выборке вопросника. Например, если пол только одной из пятнадцати самок был ошибочно классифицирован как мужской, то истинное соотношение полов 105 было бы записано как 119,6. Возможное включение шиншилл с хромосомными аномалиями, в частности половых хромосом, вносит дополнительный усложняющий фактор. Данные опроса были проверены на возможность того, что указанное соотношение полов может варьироваться в зависимости от опыта заводчика. 47 опросников вернулись с ранчо, работающих менее четырех лет. Соотношение полов среди 2467 живорожденных составило 113,78 (95% доверительный интервал от 105,14 до 123,17). Остальные двадцать два опросника были получены с ранчо старше 4 лет и показали соотношение полов среди 8197 живорождений 120,82 (95% доверительный интервал от 115,69 до 126,21). Как из этих данных, так и из данных, представленных в таблице 1, видно, что соотношение полов шиншилл имеет тенденцию быть высоким, независимо от критериев выбора подгруппы из общего количества заполненных опросных листов. Кроме того, сходство двух параметров в настоящем исследовании (соотношения полов и размера помета живорожденных), с параметрами более раннего анкетного опроса (Hillemann & Tibbitts, 1957) усиливает достоверность данных настоящего анкетирования, хотя вероятность того, что имеется подобное смещение нельзя исключить из обоих исследований.
Важный момент касается обоснованности исключения данных, предоставленных в анкете, согласно которой соотношение полов было 36,5 среди 1286 живорожденных; включение этих данных дало бы общее соотношение полов 105,75. Значительная степень неоднородности между колониями, вызванная рассмотрением этих данных, сама по себе является достаточным основанием для их исключения. Кроме того, наличие двух других аномальных особенностей серьезно ставит под сомнение достоверность данных, представленных в этом отчете. Заявленный размер помета живорожденных, равный 3,93, почти вдвое превышает значение 2,02, указанное в остальных анкетах. Точно так же это намного превышает средний показатель 1,88, полученный Hillemann & Tibbitts (1957). Заявленный коэффициент мертворождения 1,6 / 1000 живорождений намного меньше, чем либо значение 104,02, полученное Hillemann & Tibbitts (1957), либо среднее значение 76,95 в остальных вопросниках настоящего исследования. Коэффициент мертворождаемости (смертность плода) у человека составляет от 11 до 19 на 1000 живорождений (Национальный центр статистики здравоохранения США, 1967).
Чрезвычайно высокое соотношение полов у шиншиллы было подтверждено в настоящем исследовании, также было показано, что это не результат каких-либо особенностей освещения, поскольку большинство пометов было принесено на ранчо, освещенных по большей части дневным светом. Возникает соблазн предварительно приписать высокое соотношение полов у шиншиллы разнице в размерах между X- и Y-хромосомами.
Благодарности
Я признателен доктору Susumu Ohno, Медицинский центр City of Hope, Дуарте, Калифорния, за предложение этого исследования, а также редакторам Empress Chinchilla за разрешение на воспроизведение опросника (текст-рис. 1). Работа была поддержана исследовательским грантом HD 00544 Службы общественного здравоохранения США, а автор - стипендиат Джона и Мэри Р. Маркл в области академической медицины.
Использованные источники
Altman, P. L. & Dittmer, D. S., Eds. (1962) Growth. Federation of American Societies for Experimental Biology, Washington.
Bacci, G. (1965) Sex determination, p. 66. Pergamon Press, New York.
Crew, F. A. E. (1952) The factors which determine sex. In: Marshall's Physiology of Reproduction, 3rd edn. Vol. II, p. 741. Ed. A. S. Parkes. Longmans Green, London.
Galton, M. (1965) Sex ratio questionnaire. Empress Chinchilla, 21(10), 17.
Galton, M., Benirschke, & Ohno, S. (1965) Sex chromosomes of the chinchilla: allocycly and duplication sequence in somatic cells and behavior in meiosis. Chromosoma, 16, 668.
Geneva Conference (1966) Standardization of procedures for chromosome studies in abortion. Bull. WldHlth Org. 34, 765; Cytogenetics, 5, 361.
Hillemann, H. H. & Tibbitts, F. D. (1957) Reproductive performance in chinchilla. NW. Sci. 31, 92.
McFeely, R. A. (1967) Chromosome abnormalities in early embryos of the pig. J. Reprod. Fert. 13, 579.
Ott, J. N. (1964) Some responses of plants and animals to variations in wavelengths of light energy. Ann. N.T. Acad. Sci. 117, 624.
Snedecor, G. W. (1956) Statistical methods applied to experiments in agriculture and biology, 5th edn., p. 227. Iowa State College Press, Ames, Iowa.
Stevenson, A. C. (1966) Sex chromatin and the sex ratio in man. In: The Sex Chromatin, p. 263. Ed. K. L. Moore. Saunders, Philadelphia.
Tietze, C. (1948) A note on the sex ratio of abortions. Hum. Biol. 20, 156.
U.S. National Center for Health Statistics (1966) Natality statistics analysis, United States, 1963. Public Health Service Publication No. 1000, Series 21, No. 8. United States Department of Health, Education, and Welfare, Washington.
U.S. National Center for Health Statistics (1967) International comparison of perinatal and infant mortality: The United States and Six West European Countries. Public Health Service Publication No. 1000, Series 3, No. 6. United States Department of Health, Education, and Welfare, Washington.
Vickers, A. D. (1967) A direct measurement of the sex-ratio in mouse blastocysts. J. Reprod. Fert. 13,375.
Майкл Гэлтон
Департамент патологий Медицинской школы Дартмута, Хановер, Нью-Гэмпшир 03755, США
(Получено 23 июня 1967 г., исправлено 7 августа 1967 г.)
Перевод: Д.В. Михайлов
Michael Galton, Chinchilla sex ratio (Соотношение полов у шиншиллы) // J. Repord. Fert. (1968) 16, 211-216