Шласка Б. и др. Идентификация последовательностей ДНК, влияющих на поведение шиншиллы (Chinchilla lanigera Molina, 1782)
Авторы: BRYGIDA ŚLASKA (1), SYLWIA NISZTUK (1), IWONA ROZEMPOLSKA-RUCIŃSKA (1), GRZEGORZ ZIĘBA (1), MAREK BABICZ (2), MAGDALENA SURDYKA (1).
1 Department of Biological Bases of Animal Production, University of Life Sciences in Lublin, Akademicka 13, 20-950 Lublin,
2 Department of Pig Breeding and Production Technology, University of Life Sciences in Lublin, Akademicka 13, 20-950 Lublin
Краткое резюме. Цель исследования состояла в определении взаимосвязи между типами поведения шиншиллы и различными изображениями электрофореграмм, полученных с использованием метода RAPD-PCR. Тест звук-движение позволил классифицировать животных как уверенных, сдержанных и равнодушных. Результаты пилотного исследования, основанные на маркерах OPA07 и OPA12, указывают на существование областей ДНК, содержащих нуклеотидные последовательности, которые определяют поведение шиншиллы. Рекомендуется провести дальнейшие анализы с целью выявления последовательностей генетического материала, которые могут определять изученные признаки. Это облегчит использование электрофореграмм в качестве дополнительного критерия, помимо информации о происхождении, при отборе животных.
Ключевые слова: шиншилла, RAPD-PCR, поведение
Введение
Одной из общих черт в селекции видов домашних животных является мягкий характер, проявляющийся в способности подавлять страх, и низкий уровень агрессивности по отношению к человеку. Как утверждают Павлова и соавт. [2007], реакции животных на экстремальное обращение и негативные эмоции имеют особое значение. При воздействии негативного раздражителя животные проявляют пассивные стратегии поведения. Наблюдения за поведением племенных шиншилл проводились в течение относительно короткого времени. Dzierżanowska-Góryń и Kowalczyk [2005] обнаружили, что содержание животных в неблагоприятных условиях окружающей среды оказывает пагубное влияние на их психику; это может привести к самотравмированию, самопогрызанию меха и снижению аппетита. Каждый тип психологических нарушений у шиншиллы может проявляться чрезмерной агрессией, сдержанностью и робостью в ответ на новый раздражитель. Поэтому агрессивные или робкие животные должны исключаться из разведения. Такие животные характеризуются более низкими темпами роста; им трудно рожать и выращивать потомство. Иногда молодые шиншиллы съедаются или травмируются матерями, подвергнутыми стрессу.
Выбор поведенческого теста является ключевым элементом в определении типа поведения шиншиллы [Ślaska and Jeżewska-Witkowska 2008a]. Вышеупомянутые результаты исследования позволяют сделать вывод, что вместо улучшения достаточно благоприятных условий окружающей среды на шиншилловых фермах, отбор наиболее благополучных животных с точки зрения репродуктивности может быть выполнен в условиях, установленных человеком. Такое управление может принести ощутимую пользу.
При разведении пушных зверей генетические маркеры используются для контроля происхождения, точного определения сводных родственников в одном помете, построения генетических карт и выявления генетической дистанции и генетических различий и сходств между различными группами особей. Путем выявления генетической изменчивости молекулярные маркеры могут предоставить ценную информацию о популяции и ее структуре, уровне дрейфа генов, филогенетических взаимосвязях, исторических биогеографических моделях, а также анализе происхождения и родства. Результаты пилотного исследования, основанного на использовании маркеров RAPD [Ślaska и Jeżewska-Witkowska 2008b], свидетельствуют о существовании областей ДНК, содержащих нуклеотидные последовательности, которые отвечают как за массу тела, так и за общий экстерьер шиншилл. Поэтому маркеры RAPD могут быть использованы в качестве дополнительного критерия для формирования генетических групп при отборе животных в разведение.
Цель исследования состояла в том, чтобы определить взаимосвязь между поведением шиншиллы и различными изображениями электрофореграмм, созданных с использованием метода RAPD.
Материалы и методы
Материал исследования состоял из шиншилл окраса стандарт (33 самки и 15 самцов), родившихся на одной из племенных ферм в восточной Польше в 2006-2007 годах. Животных содержали в стандартных клетках с подстилкой, освещенных искусственным светом при 12-часовом дневном световом режиме и при температуре ок. 18 °C (+/- 2 °C). Для оценки характера шиншилл был проведен тест звук-движение, описанный Ślaska и Jeżewska-Witkowska [2008a]. Выбор теста из нескольких поведенческих тестов, применяемых у шиншилл, был обусловлен результатами исследования по репродуктивному благополучию шиншилл [Ślaska and Jeżewska-Witkowska 2008a].
Наблюдения проводились между 4 и 8 часами при дневном свете человеком, неизвестным шиншиллам, так как владелец фермы проводил санитарную и ветеринарную обработку. Это вызывало специфичное поведение отдельных животных по отношению к их заводчику.
Основываясь на наблюдениях за поведением шиншилл в поведенческом тесте, самки, которые проявляли похожий тип реакции, классифицировали по следующим группам:
- Спокойные животные-исследователи - те, кого не пугал звук открывающейся двери; они немедленно приближались к наблюдателю и выскакивали из клетки в течение 20 секунд; у них не было никакой реакции, которая могла бы подразумевать чувство беспокойства по отношению к незнакомцу;
- Осторожные животные-исследователи - эта группа состояла из животных, которые проявляли интерес в течение 60 секунд после начала наблюдения, то есть особей, которые приблизились к объекту или открытой дверце в течение 20-60 секунд; эти особи подходили к дверце, но не высовывались из нее; они были полностью бдительны и готовы убежать от возможного внешнего раздражителя;
- Безразличные животные - те, которые не реагировали на раздражитель звук-движение; они обычно сидели на полке или в углу клетки и не проявляли особого интереса к внешней среде.
ДНК выделяли с использованием QIAamp DNA Blood Mini Kit (QIAGEN) из волосяных фолликулов шиншилл, участвовавших в эксперименте. Реакция ПЦР сопровождалась количественной и качественной оценкой образцов ДНК. Реакционная смесь (объем образца 30 мкл) содержала 80 нг ДНК; 3 мкл ПЦР-буфера, 4,2 мкл Q-раствора, 200 мкМ каждого нуклеотида, 0,2 мкМ произвольного праймера, 25 мМ MgCl2, 1 ед. ДНК полимеразы Taq. Для реакции амплификации использовали два праймера: OPA07 (5 ’→ 3’GAAACGGGTG) и OPA12 (5’ → 3’TCGGCGATAG) (Proligo Primers and Probes).
Маркеры были выбраны на основе их полиморфизма, описанного в исследовании Ślaska et al. [2008]. Реакция амплификации (PTC-225 DNA Engine Tetrad thermocycler) заключалась в начальной денатурации (94 °С, 5 мин); 46 циклов: денатурация (94 °С, 1 мин.), связывание праймера (36 °С, 2 мин.), удлинение цепочек ДНК (72 °С, 1 мин.); терминальное удлинение праймеров (72 °С, 10 мин.) и охлаждение до темп. 4 °C. Продукты RAPD-PCR фракционировали в 2%-м агарозном геле (с использованием загрузочного раствора бромфенола синего). Гели были проанализированы в УФ-свете (Transiluminator) и архивированы. В качестве маркеров размера фрагмента ДНК были использованы маркер длины ДНК GeneRuler 50 п.н. или маркер длины ДНК GeneRuler100 п.н. (Fermentas).
Вероятность обнаружения специфических фрагментов ДНК, связанных с характером особей, идентифицированных с помощью теста звука-движения, была подтверждена с помощью однофакторного дисперсионного анализа [SAS Institute INC 2000]. На основании наличия (1) или отсутствия (0) полосы каждого из маркеров, исследованных в агарозном геле, была определена вероятность его присутствия у животных, проявляющих определенные типы поведения. Это было сделано с целью обнаружения полос, которые могут содержать последовательности ДНК, определяющие поведение шиншиллы. Амплифицированные фрагменты ДНК длиной 1050, 1200 и 1500 пар нуклеотидов (в случае OPA07) и 600 п.н. (OPA12) были исключены из анализа, поскольку они были обнаружены у всех протестированных животных.
Эти пилотные исследования, направленные на обнаружение специфичных полос ДНК, которые могут содержать гены или регуляторные области генов, влияющих на определенные типы поведения животных, являются предварительными. Они будут стимулировать дальнейшие исследования, сфокусированные на SCAR - последовательностях, характеризующих амплифицированные области - маркеры, происходящие от маркеров RAPD [Cushwa and Medrano 1996, Gu et al. 1999]. Они выявят участки ДНК, которые могут содержать вышеупомянутые нуклеотидные последовательности.
Результаты и обсуждение
Полиморфизм RAPD возникает в результате наличия или отсутствия определенных полос, генерируемых праймером, причиной этого может быть различие нуклеотидной последовательности в области связывания праймера. Мутация только с одной стороны нуклеотида приводит к отсутствию амплификации. Присутствие или отсутствие результата также может быть вызвано вставкой или удалением фрагмента между двумя консервативными сторонами связывания праймера. Исследования генома Chinchilla lanigera с использованием маркеров RAPD-PCR ранее проводились у шиншилл [Hidas et al. 2002, Ślaska and Jeżewska-Witkowska 2008b, Ślaska et al. 2008]. Ślaska et al. [2008] оценивали пригодность маркеров RAPD-PCR для генетического анализа у шиншилл; среди 10 генетических маркеров наиболее полиморфные фрагменты ДНК были получены, среди прочего, благодаря маркерам OPA07 и OPA12.
В исследуемой группе шиншилл спокойные животные-исследователи составляли 35,42% (47,26% самцов, 52,94% самок), осторожные животные-исследователи - 35,42% (29,41% самцов, 70,59% самок) и безразличные животные - 29,17% (14,29% самцов, 85,71% самок) из всех проверенных особей. Среди животных, наблюдаемых на племенной ферме, не было зарегистрировано агрессивного поведения.
Отсутствие шиншилл, характеризующихся агрессивным поведением, на экспериментальной ферме может быть объяснено тем фактом, что, в отличие от животных с умеренным темпераментом, агрессивные особи исключаются из разведения из-за их более низких темпов роста и более низких показателей репродуктивности и, в частности, выращивания потомства [Dzierżanowska-Góryń and Kowalczyk 2005, Gacek 2002].
Исследования типа поведения шиншиллы при помощи маркеров OPA07 и OPA12 носят экспериментальный характер. На основании проведенного анализа вероятность присутствия определенных полос, связанных с типом поведения шиншиллы, была определена с помощью теста звука-движения и маркеров OPA07 (Рис. 1, Табл. 1) и OPA12 (Рис. 2, Табл. 2).
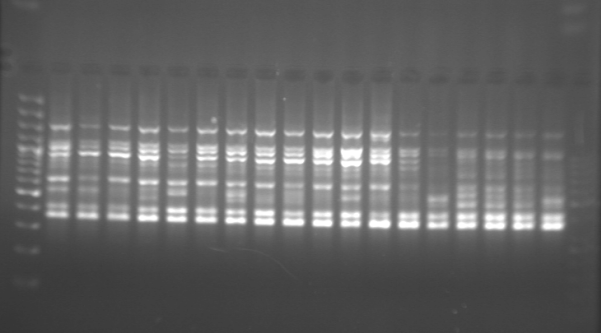
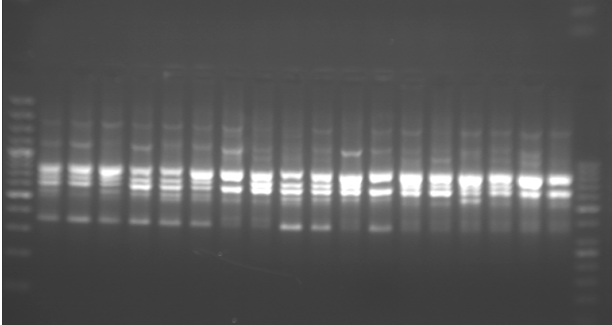
Исследование не включает разделение животных по мужской и женской группам в рамках определенных типов поведения, так как никаких существенных различий в вероятности присутствия индивидуальных полос в этих группах животных не наблюдалось. Более того, в случае обоих использованных при анализе маркеров ни одна из полученных полос не была характерна исключительно для одного пола.
|
Размер амплифицированных фрагментов ДНК (п.н.) ** |
Тип поведения |
|||||
| Спокойные животные-исследователи | Осторожные животные-исселдователи | Безразличные животные | ||||
|
P* |
se |
P* |
se |
P* |
se |
|
|
250 |
0.118 |
0.059 |
0.000 |
0.059 |
0.071 |
0.065 |
|
300 |
0.412 |
0.120 |
0.294 |
0.120 |
0.429 |
0.132 |
|
350 |
0.059b |
0.101 |
0.294 |
0.101 |
0.429 a |
0.111 |
|
400 |
1.000 |
0.047 |
1.000 |
0.047 |
0.857 |
0.052 |
|
500 |
0.471 |
0.115 |
0.765 |
0.115 |
0.714 |
0.126 |
|
550 |
0.000 b |
0.065 |
0.059 |
0.065 |
0.214 a |
0.072 |
|
600 |
0.470 |
0.121 |
0.706 |
0.121 |
0.500 |
0.133 |
|
700 |
0.941 a |
0.099 |
0.765 |
0.099 |
0.500b |
0.109 |
|
800 |
0.941 a |
0.097 |
0.765 |
0.097 |
0.643 b |
0.107 |
|
900 |
0.765 |
0.110 |
0.765 |
0.110 |
0.643 |
0.121 |
a, b - Значения в строках, помеченных различными буквами, значимо различаются при p ≤ 0,05** - приблизительные значения
Что касается праймера OPA07 (Табл. 1), особое внимание следует уделить полосам 350, 550, 700 и 800 п.н. Вероятность появления полос длиной 700 и 800 п.н. была значительно выше в группе спокойных животных-исследователей, чем у безразличных шиншилл. Статистически значимые различия относительно вероятности присутствия полос 350 и 550 п.н. были также обнаружены между группами, упомянутыми выше. У безразличных шиншилл вероятность присутствия вышеупомянутых фрагментов была выше, чем у других групп животных. Из этого, возможно, следует, что исследованные фрагменты ДНК содержат нуклеотидные последовательности, которые могут оказывать влияние на тип поведения шиншиллы.
При анализе данных, представленных в таблице 2, следует обратить внимание на значимые различия, касающиеся наличия фрагментов ДНК в конкретных группах шиншилл, имеющих различное поведение. По сравнению с другими животными полоса 400 п.н. показала наибольшую вероятность присутствия в группе спокойных животных-исследователей. Фрагменты ДНК размером 800 и 1750 п.н. показали наибольшую вероятность появления в группе безразличных шиншилл.
|
Размер амплифицированных фрагментов ДНК (п.н.) ** |
Тип поведения |
|||||
|
Спокойные животные-исследователи |
Осторожные животные-исселдователи |
Безразличные животные |
||||
|
P* |
se |
P* |
se |
P* |
se |
|
|
350 |
0.709 |
0.100 |
0.882 |
0.100 |
0.786 |
0.110 |
|
400 |
0.647 |
0.119 |
0.471 |
0.119 |
0.286b |
0.131 |
|
500 |
0.882 |
0.081 |
0.941 |
0.081 |
0.786 |
0.089 |
|
550 |
0.588 |
0.116 |
0.765 |
0.116 |
0.643 |
0.128 |
|
700 |
0.588 |
0.114 |
0.706 |
0.114 |
0.786 |
0.126 |
|
800 |
0.647b |
0.106 |
0.647b |
0.106 |
0.929a |
0.117 |
|
900 |
0.353 |
0.101 |
0.235 |
0.101 |
0.071 |
0.111 |
|
1050 |
0.471 |
0.115 |
0.294 |
0.115 |
0.214 |
0.126 |
|
1200 |
0.412 |
0.124 |
0.471 |
0.124 |
0.571 |
0.136 |
|
1500 |
0.235 |
0.106 |
0.176 |
0.106 |
0.357 |
0.117 |
|
1750 |
0.000b |
0.086 |
0.176 |
0.086 |
0.357a |
0.094 |
|
2000 |
0.118 |
0.082 |
0.176 |
0.082 |
0.071 |
0.090 |
Результаты, полученные другими авторами [Dzierżanowska-Góryń and Kowalczyk 2005, Rozempolska-Rucińska et al. 2007], показывают, что заводчики должны проводить селекцию с целью получения спокойных животных-исследователей. Это связано с тем, что самки с умеренным темпераментом характеризуются более высокой скоростью размножения по сравнению с робкими или агрессивными. Однако, по мнению других авторов [Gacek 2002, Ślaska and Jeżewska-Witkowska 2008a], самки с умеренным темпераментом не желательны для разведения из-за более низкой плодовитости и более низкой продуктивности при выращивании потомства.
Исследования, проведенные Gu et al. [1999] продемонстрировали, что одна полоса определенного размера, полученная реакцией RAPD, может состоять из более чем одного фрагмента ДНК того же размера, но с другой нуклеотидной последовательностью. Однако Espinasa и Borowsky [1998] сообщили о необходимости подтвердить гомологию полос, полученных при исследованиях филогенетически удаленных видов. В настоящем исследовании подтверждение гомологии представляется ненужным, так как в исследовании участвовал один вид. Следовательно, предположение о том, что анонимные фрагменты ДНК одинакового размера, полученные в профилях отдельных животных, представляют собой гомологичные локусы, является обоснованным.
Предыдущие исследования групп шиншилл [Ślaska and Jeżewska-Witkowska 2008b] позволили выявить взаимосвязь между массой тела и общим экстерьером, а также получить различные изображения электрофореграмм, с помощью метода RAPD. Результаты предполагают наличие последовательностей ДНК в определенных полосах, определяющих количественные признаки.
На основании оценки успешности размножения шиншилл [Ślaska and Jeżewska-Witkowska 2008a] был сделан вывод о том, что самки, классифицированные по критерию звук-движение как осторожные животные-исследователи, характеризовались самой высокой плодовитостью и количеством отлученного от матери потомства на один помет. Настоящие результаты (табл. 1) показывают, что последовательность 250 п.н. (OPA07) у осторожных шиншилл отсутствовала. Вероятность присутствия полосы размером 600 п.н. была также выше в упомянутой группе по сравнению с таковой у спокойных животных-исследователей и безразличных животных. В случае маркера OPA12 (табл. 2) существенные различия в присутствии фрагментов ДНК размером 400, 800 и 1750 п.н. указывают на необходимость продолжения исследования. Дальнейший анализ (SCAR) фрагментов ДНК рекомендуется в связи с возможной идентификацией содержащихся в них последовательностей. Не исключено, что внутри каждого фрагмента ДНК анализируемых маркеров имеются нуклеотидные последовательности, которые могут существенно влиять на тип поведения шиншиллы.
Заключение
- Отдельные фрагменты ДНК маркеров OPA07 и OPA12 следует использовать для дальнейших анализов, направленных на идентификацию содержащихся в них нуклеотидных последовательностей.
- Не исключено, что после дополнительных генетических анализов маркеры OPA 07 и OPA12 будут использоваться в качестве дополнительного критерия при создании генетических групп животных согласно их поведению.
- Результаты исследования могут способствовать использованию в будущем отдельных фрагментов ДНК в качестве дополнения к данным родословной для генетической сохранности отдельных типов поведения.
Использованные источники
Cushwa W.T., Medrano J.F., 1996. Applications of the randomly amplified polymorphic DNA (RAPD) assay for genetic analysis of livestock species. Anim. Biotechnol. 7, 11–31.
Dzierżanowska-Góryń D., Kowalczyk M., 2005. Wstępne badania temperamentu szynszyli. Rocz. Nauk. PTZ, Supl. 1, 97–103.
Espinasa L., Borowsky R., 1998. Evolutionary Divergence of AP-PCR (RAPD) Patterns. Mol. Biol. Evol. 15, 408–414.
Gacek L., 2002. Test empatyczny – nowy test behawioralny dla lisów polarnych. Przegl. Hod. 5, 4–6.
Gu W., Post M., Aguirre G.D., Ray K., 1999. Individual DNA bands obtained by RAPD analysis of canine genomic DNA often contain multiple DNA sequences. J. Hered. 90, 96–98.
Hidas A., Edvi M.E., Potháczky L., Tóth Á., 2002. RAPD polymorphism in chinchilla breeding stocks. Proceedings of the 7th World Congress on Genetics Applied to Livestock Production,
Montpellier, France, August, 2002. Session 12, 2002 pp. 0–3.
Pavlova I.V., Levshina I.P., Vanetsian G.L., Shuikin N.N., Zyabitseva E.A., (2007). Behavior and measures of respiration in rabbits differing in terms of movement activity in ano pen field. Neurosci. Behav. Physiol. 37, 33–41.
Rozempolska-Rucińska I., Jeżewska G., Mielnik A., 2007. Wyniki rozrodu samic i odchowu młodych szynszyli w zależności od temperamentu matki. Rocz. Nauk. PTZ 3, 283–294.
SAS Institute Inc., 2000. SAS User’s Guide. Version 8.0 Edition, SAS Institute Inc., Cary.
Ślaska B., Jeżewska-Witkowska G., 2008a. Wykorzystanie testów behawioralnych do oceny dobrostanu reprodukcyjnego szynszyli (Chinchilla lanigera Molina, 1782). Rocz. Nauk. PTZ 4, 323–335.
Ślaska B., Jeżewska-Witkowska G., 2008b. Ocena związku pomiędzy cechami użytkowymi szynszyli (Chinchilla lanigera Molina, 1782) a obrazem DNA. Folia Univ. Agric. Stetin., Agric., Aliment., Pisc., Zootech. 266, 177–184.
Ślaska B., Jeżewska G., Zięba G., 2008. The utility of RAPD-PCR markers for evaluation genetic differentiation of chinchilla (Chinchilla lanigera Molina, 1782). 13 Genetic Days, September, 39–43.
B. Ślaska, S. Nisztuk, I. Rozempolska-Rucińska, G. Zięba, M. Babicz, M. Surdyka. Identification of DNA sequences affecting chinchilla (Chinchilla lanigera Molina, 1782) behaviour (Идентификация последовательностей ДНК, влияющих на поведение шиншиллы (Chinchilla lanigera Molina, 1782)) // Annales Universitatis Mariae Curie-Skłodowska. Sectio EE: Zootechnica, 2012, T. 30, №2, p.63-70.