Ванжонак В.Дж., Джонсон Х.Д. Взаимосвязь функции щитовидной железы и надпочечников с самопогрызанием меха у шиншилл
Тип статьи:
перевод
Вид статьи:
экспериментальная статья
Аннотация
-
Шиншиллы, выгрызавшие мех, продемонстрировали повышенную активность щитовидной железы (P<0,05), увеличенную функцию надпочечников (P<0,05) и пониженную ректальную температуру (P<0,01).
-
Это исследование показало, что повышение эндокринной активности может быть связано с утратой изоляции в областях, где мех выгрызен.
-
Однако, возможно, что эндокринная активность может быть стимулом к самопогрызанию меха.
-
Также возможно самопогрызание меха может начаться или же его распространенность увеличивается при наличии "неспецифических стрессоров".
Введение
Установлено, что около 30 процентов шиншилл (Chinchilla lanigera) имеют странную жевательную привычку, при которой животные выгрызают мех (Rees, 1962). Имеется мало фактических данных о причинах самопогрызания меха.
Чтобы обеспечить физиологическую основу для понимания самопогрызания меха и получить данные о видах, которым специалисты по сравнительной физиологии или экофизиологи уделяют мало внимания, в лаборатории было начато исследование, в котором были измерены функция надпочечников и щитовидной железы и потребление кислорода у шиншилл в норме и страдающих самопогрызанием меха.
Материалы и методы
В данном исследовании было использовано двенадцать шиншилл (Chinchilla lanigera), шесть здоровых и шесть с самопогрызанием. Все животные были сопоставимы по массе тела и возрасту. Животные были получены от поставщика (Pioneer Chinchilla Ranch, 148 Linda Vista Avenue, North Haledon, New Jersey) в возрасте 8 месяцев и сразу разделены на две группы, нормальную и с самопогрызанием. Животных содержали при термонейтральной температуре 22 °C (Drozdz & Gorecki, 1967) и помещали по отдельности в подвесные клетки 10 x 7 x 7 дюймов с проволочной сеткой по бокам, сверху и снизу и дном из твердого оцинкованного железа. Корм «Purina chinchilla chow» и вода были в свободном доступе. Условия содержания оставались одинаковыми на протяжении всего исследования.
Измерения проводились в обеих группах в возрасте 12 и 18 месяцев. Для определения скорости высвобождения гормонального 131I из щитовидной железы использовали метод Johnson et al. (1964). Была проведена подкожная инъекция в дозе 2.5 UC 131I, учет функции щитовидной железы проводили дважды в день, через 24-84 часа после инъекции. Наклон полулогарифмической регрессии, экспонента k, был использован в качестве одного из показателей активности щитовидной железы (Johnson & Ragsdale, 1960).
Для получения дополнительной оценки функции щитовидной железы была определена скорость поглощения 131I в течение 24 часов и уровни секреции щитовидной железы (TSR) по методу Grosvenor & Turner (1958), в котором используется инъекция увеличивающихся доз экзогенного L-T4 для подавления секреции TSH и, таким образом, подавления высвобождения щитовидной железой 131I. Животным вводили подкожно 10 UC 131I до инъекции тироксина. Ежедневные инъекции 1,6 мг метимазола / 100 г массы тела (м.т.) предотвращали рециркуляцию 131I из метаболизированного тироксина. TSR оценивался в возрасте 18 месяцев в обеих группах.
Глюкокортикоиды в плазме измеряли у двух экземпляров по методу Guillemin et al. (1959), за исключением того, что метиленхлорид использовали в качестве экстракционного растворителя, а серную кислоту с содержанием этанола использовали в качестве флуоресцентного реагента (Mattingly, 1962).
Потребление кислорода определялось по методу непрямой замкнутой системы (Kibler, 1967). Температуру тела измеряли с помощью специально откалиброванного ректального зонда с ограниченным диапазоном действия (Kibler & Johnson, 1967). Ежедневное потребление корма и масса тела измерялись у всех животных во время физиологических измерений. Измерение отношения массы к площади меха (мг/см2) определяли путем удаления непрерывно растущей области меха в области бедра и спины с помощью машинки для стрижки.
Результаты
При использовании показателей выделения 131I щитовидной железой (k) в качестве меры активности щитовидной железы, у животных, выгрызающих мех, было получено значительно более высокое (P<0,001) значение k (8,44), чем у контрольной группы без самопогрызания (5,73) в возрасте 12 месяцев (таблица 1). Значение k у шиншилл с самопогрызанием было значительно выше в возрасте 18 месяцев (P<0,001). В возрасте 12 месяцев измерение поглощения 131I у шиншилл с самопогрызанием было значительно выше (P <0,01), чем у нормальных особей.
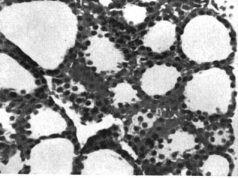
Чтобы подтвердить эти значимые результаты активности щитовидной железы, были определены показатели секреции щитовидной железы (TSR) в возрасте 18 месяцев (таблица 1). Значимо более высокий уровень TSR (3,07 / мкг L-T4 / 100 г (массы тела в сутки) был получен у шиншилл с самопогрызанием (по сравнению с нормальным значением 2,35). Рисунок 1 (а) демонстрирует репрезентативные гистологические наблюдения пяти нормальных и шести животных с самопогрызанием меха. Группа с самопогрызанием меха имела более высокий уровень клеток цилиндрического секреторного эпителия и меньше коллоидов, что свидетельствует о более высоком функциональном состоянии железы (Turner, 1967).
Животные, выгрызавшие мех, имели значительно более высокие (P<0,001) уровни кортикостерона в плазме (0,451 мкг/мл по сравнению с 0,253 мкг/мл для нормальных животных) (таблица 1). На рис. 1 (b) показаны репрезентативные гистологические наблюдения шести нормальных и шести животных с самопогрызанием. В группе с самопогрызанием наблюдается чрезмерная гипертрофия надпочечников. Такая адренокортикальная гипертрофия с повышенным уровнем кортикостерона в плазме является убедительным доказательством более высокого функционального состояния коры надпочечников в группе с самопогрызанием (Turner, 1967).
Измерение массы меха на единицу площади показало 50-процентную массу меха (мг/см2) в областях самопогрызания, по сравнению с нормальными участками (таблица 1).
Средняя ректальная температура в группе с самопогрызанием составляла 35,96 °C в возрасте 12 месяцев и 35,92 °C в возрасте 18 месяцев. В обоих возрастах она была значимо более низкой (P<0,01), чем у нормальных особей (36,75 и 36,46 °C соответственно).

Рисунок 2(а). Общий внешний вид нормальной шиншиллы
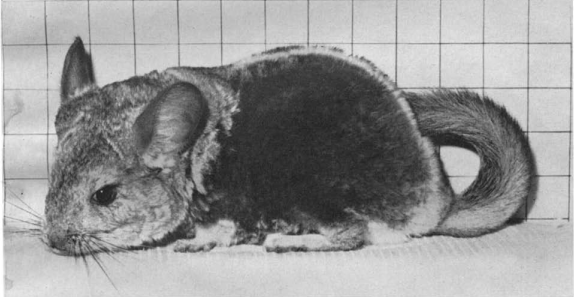
Рисунок 2(b). Общий внешний вид шиншиллы, страдающей самопогрызанием меха
| Животное с самопогрызанием меха | Нормальное животное |
Щитовидная железа (х440) |
|
 |
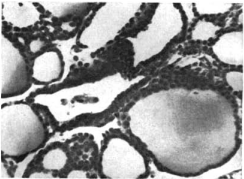 |
| Рисунок 1 (а) | |
Надпочечник (х40) |
|
 |
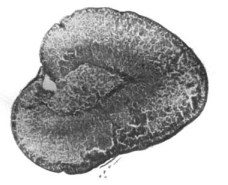 |
| Рисунок 1 (b) | |
Рисунок 1. (а) Гистология типичной щитовидной железы у шиншилл с самопогрызанием меха и нормальных шиншилл. (b) Гистология типичного надпочечника у шиншилл с самопогрызанием меха и нормальных шиншилл.
Таблица 1. Физиологические измерения шиншилл с самопогрызанием меха и нормальных в возрасте 12 и 18 месяцев.
|
|
Значение для нормальных животных
|
Кол-во
|
Ст. откл.
|
Значение для
шиншилл с
самопогрызанием
|
Кол-во
|
Ст. откл.
|
t-критерий,
ур. знач.
|
%
изменений
|
|
Возраст 12 месяцев
|
||||||||
|
Выделение I131 (k x 10-3)
|
5,73
|
5
|
±0.4
|
8,44
|
6
|
±0.8
|
P<0.001
|
+52.5
|
|
Поглощение I131 (24 ч)
|
6,45
|
6
|
±0.3
|
9,22
|
5
|
±0.5
|
P<0.01
|
+42.9
|
|
Ректальная температура (°С)
|
36,75
|
5
|
±0.1
|
35,96
|
6
|
±0.1
|
P<0.01
|
-2.04
|
|
VO2 (мл/ч)
|
437,62
|
4
|
±57.8
|
431,07
|
4
|
±5.8
|
Н.з.
|
-1.4
|
|
VO2 / массу тела
|
1,13
|
5
|
±0.1
|
1,03
|
4
|
±0.1
|
Н.з.
|
-8.8
|
|
Масса тела (г)
|
384,6
|
5
|
±24.0
|
415
|
6
|
±9.3
|
Н.з.
|
+7.9
|
|
Потребление пищи (г)
|
15,37
|
5
|
±1.1
|
15,42
|
6
|
±0.9
|
Н.з.
|
+0.3
|
|
Возраст 18 месяцев
|
||||||||
|
Скорость выделения I131 (kx10-3)
|
4,46
|
5
|
±0.8
|
9,40
|
7
|
±0.6
|
P<0.001
|
+110
|
|
Поглощение I131 (24 ч) (мкг/100 г м.т. в сутки)
|
6,8
|
6
|
±0.4
|
11,8
|
7
|
±0.8
|
P<0.05
|
+57.6
|
|
TSR
|
2,35
|
10
|
±0.07
|
3,07
|
7
|
±0.06
|
P<0.001
|
+30.6
|
|
Кортикостерон в плазме (мкг/мл)
|
0,253
|
7
|
±0.062
|
0,451
|
7
|
±0.068
|
P<0.001
|
+70.1
|
|
Масса меха (мг/см2)
|
|
|
|
|
|
|
|
|
|
На бедре
|
106
|
6
|
±10.2
|
49
|
7
|
±7.0
|
P<0.01
|
-53.7
|
|
Сзади
|
109
|
9
|
±8.7
|
44
|
4
|
±6.7
|
P<0.01
|
-59.6
|
|
Ректальная температура (°С)
|
36,46
|
6
|
±0.2
|
35,92
|
7
|
±0.1
|
P<0.01
|
-1.5
|
|
VO2 (мл/ч)
|
361,9
|
8
|
±75.8
|
349,8
|
6
|
±32.8
|
Н.з.
|
-3.3
|
|
VO2 / массу тела
|
0,863
|
5
|
±0.08
|
0,82
|
6
|
±0.09
|
Н.з.
|
-4.9
|
|
VO2 / массу тела на °С
|
0,021
|
6
|
±0.001
|
0,026
|
6
|
±0.001
|
P<0.05
|
+25.0
|
|
Масса тела (г)
|
410,8
|
5
|
±12.9
|
455,86
|
7
|
±33.5
|
Н.з.
|
+10.9
|
|
Потребление пищи (г)
|
16,22
|
5
|
±1.2
|
16,52
|
6
|
±0.9
|
Н.з.
|
+1.8
|
Н.з. – не значимо
VO2 на животное и VO2 на г массы тела существенно не различались между двумя группами; однако, если скорость метаболизма была выражена как VO2/массу тела на °С (ректальная температура), у особей, имевших самопогрызание меха в возрастной группе 18 месяцев, (P<0,05) уровень метаболизма был несколько более высоким (0,026 против 0,021) (таблица 1).
Сравнение средней массы тела контрольной группы и шиншилл с самопогрызанием не показало существенных различий в возрасте 12 или 18 месяцев (таблица 1), также не наблюдалось значимых различий в потреблении корма.
Обсуждение
Данное исследование указывает на то, что шиншиллы, страдающие самопогрызанием, имеют повышенную активность щитовидной железы, повышенный уровень кортикостерона в плазме и пониженную ректальную температуру по сравнению с нормальными шиншиллами. Ни потребление корма, ни VO2 не продемонстрировали значимых различий между двумя группами в возрасте 12 и 18 месяцев.
Возможным объяснением отсутствия значительного увеличения VO2 / массу тела у шиншилл с самопогрызанием, является то, что увеличение активности щитовидной железы не давало пропорционального увеличения метаболического или калоригенного уровня у животного из-за большей скорости деградации тироксина и/или повышенной потери кала (Blahd, 1971) или увеличения тироксиносвязующей способности белка (Turner, 1967).
Из данных в Таблице 1 индекс изоляции можно рассчитать по формуле (индекс изоляции = температура тела-температура окружающей среды / VO2) согласно Hart & Heroux (1955). Этот индекс показывает, что теплоизоляция у шиншилл с самопогрызанием будет ниже. Группа, которая страдала самопогрызанием, имела на 50% меньше шерсти (вес / площадь) по сравнению с поврежденными выгрызанием участками, что объясняло бы меньшую изоляцию (рис. 2а, б). Поскольку группа с самопогрызанием имела меньшую изоляцию и, по-видимому, более низкую температуру кожи, это могло вызывать повышенное срабатывание периферических рецепторов холода (Hensel, 1968). Кроме того, пониженная температура тела, действующая непосредственно на центральную нервную систему, может обеспечить стимуляцию глубоких процессов организма, а также периферическое кожное раздражение через гипоталамус (CRF, TRF) и переднюю долю гипофиза (ACTH, TSH) для повышения активности щитовидной железы и коры надпочечников (Hensel, 1968).
Во время исследования мы наблюдали двух нормальных шиншилл с нормальной активностью щитовидной железы, которые начали выгрызать мех и остались таковыми на протяжении всего исследования. После того, как они начали выгрызать мех, активность их щитовидных желез стала выше, чем в исходной группе шиншилл с самопогрызанием (рис. 3). Эти два животных также показали снижение ректальной температуры.
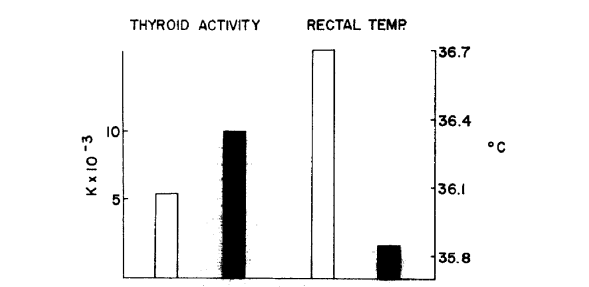
Рисунок 3. Сравнение значений двух нормальных животных (белый столбик), которые стали заниматься самопогрызанием (черный столбик), в период между 12 и 18 месяцами измерения.
Прим. пер. к рис. 3: Thyroid activity – активность щитовидной железы, Rectal temp. – ректальная температура.
Для дополнения связи щитовидной железы с изменениями термоизоляции, стоит заметить, что линька контролируется несколькими эндокринными факторами, одним из которых является щитовидная железа (Sturkie, 1965). Существуют также данные о том, что у куропаток, имеющих скудное оперение, уровень активности щитовидной железы относительно высок (Benedict et al., 1932). Фризские куры также имеют субнормальные ректальные температуры (Sturkie, 1965). Две шиншиллы находились в процессе линьки как раз перед тем, как начали заниматься самопогрызанием, это общее наблюдение у заводчиков шиншилл.
Кеннеди (1952) сообщал, что при температуре окружающей среды 21-32 °C самопогрызание меха развилось практически у всех шиншилл в группе. Когда ту же группу подвергали воздействию температуры окружающей среды, не превышающей 10 °С, самопогрызание прекращалось. Возможно, что линька меха у шиншилл также вызвана более высокой активностью щитовидной железы. В случаях самопогрызания меха активность щитовидной железы остается повышенной, и животные, вероятно, выгрызают свой мех, чтобы увеличить теплоотдачу за счет снижения теплоизоляции, и таким образом предотвратить значительное увеличение VO2.
Разные заводчики шиншилл и фермеры (Kennedy, 1952; Houston & Prestwich, 1962) отмечали, что шиншиллы с самопогрызанием имеют тенденцию быть более «нервными и возбудимыми» и что внезапные изменения окружающей среды (перевозка, изменение рациона питания, частые и резкие изменения рутинного образа жизни) могут привести к увеличению самопогрызания. Авторы также отметили, что в поголовьях, в которых отсутствуют условия рационального регулирования содержания (антисанитарные условия, плохое питание и т. д.), частота самопогрызания меха обычно выше. Koch (1968) сообщил, что в случаях хронического раздражения кишечника и адгезии частота самопогрызания меха увеличивается, поэтому возможно возникновение самопогрызания или увеличение его частоты в результате «неспецифического стресса» у животного. Повышенные уровни кортикостерона в плазме и гипертрофия надпочечников, о которых сообщается в данном исследовании, могут быть вызваны «неспецифическими стрессорами», действующими через гипоталамус, что стимулирует секрецию рилизинг-фактора кортикотропина, инициирующего высвобождение АКТГ, что в свою очередь вызывает трофические реакции коры надпочечников (Хензель, 1968).
Поскольку самопогрызание меха не свойственно шиншиллам и встречается у других фермерских пушных зверей (Houstin & Prestwich, 1962), необходимо провести дальнейшие исследования, чтобы определить роль эндокринов в феномене самопогрызания меха.
Заключение
Шиншиллы, которые страдают самопогрызанием меха, имеют значимо повышенную активность щитовидной железы и надпочечников и пониженную температуру тела. Это исследование предполагает, что повышенная эндокринная активность может быть связана с утратой изоляции в областях с поврежденным мехом; либо же повышенная эндокринная активность может быть стимулом для самопогрызания меха; или самопогрызание меха может быть вызвано или его частота увеличивается из-за "неспецифических стрессоров" среды обитания животных.
Источники
BENEDICT F. G., LANDAUER F. G. 8: Fox E. (1932) The physiology of normal and frizzlefowl, with special reference to the basal metabolism. Univ. Con. (Storrs). Agr. Sta.Bull. 1'77, 15.
BLAHD W. H. (1971) Nuclear Medicine, p. 186. McGraw-Hill, New York.
DROZDZ A. & GORECKI A. (1967) Oxygen consumption and heat production in chinchillas. ACTA Theriol. 12, 81-86.
GROSVENOR C. E. 8: TURNER C. W. (1958) Effect of lactation upon thyroid secretion rate in the rat. Pros. Soc. Exp. Biol. Med’. 99, 517-519.
GUILLEMIN R., CLAYTON G. W., LIPSCOMB H. S. 8: SMITH J. D. (1959) Fluorometric measurement of rat plasma and adrenal corticosterone concentration. j’. Lab. clin. Med. 53, 830-832.
HART J. S. 8: HEROUX O. (1955) Exercise and temperature regulation in lemmings andrabbits. Can. IBiochem. Physiol. 33, 42841-35.
HENSEL H. (1968) Adaption to Cold. In Adaptation of Domestic Animals (Edited by HAFEZ E. S. E.). Lea 8c Febiger, Philadelphia.
HOUSTON J. W. 8: Pnmrwrcrr ]. P. (1962) Chinchilla Care, p. 311. Bordon, Los Angeles.
JOHNSON H. D. 8: KIBLER H. H. (1964-) The influence of ambient temperature of 9°C and 28°C on thyroid function of rats during growth and aging. Gerontologia 9, 18-27.
JOHNSON H. D. 8: RAGSDALE A. C. (1960) The effect of rising environmental temperatures(35-—95°F) on thyroid I131 release rate of Holstein, Brown Swiss and Iersey heifers. J Agric. Sci. 54, 421-426.
KENNEDY A. H. (1952) Chinchilla Diseases and Ailments, p. 259. Fur Trade Journal of Canada, Toronto.
KIBLER H. H. 8: JOHNSON H. D. (1967) Growth and metabolism in rats as affected by dietary restriction, environmental temperature and endocrine therapy. Growth 31, 205-216.
KOCH B. (1968) Diseases and ailments. Emp. Chin. Ranch Handbook 10, 3-16.
MATTINGLY D. (1962) A simple fluorometric method for the estimation of free 11-hydroxy-corticoids in human plasma. J. Clin. Path. 915, 374-379.
REES R. G. (1962) Fur-chewing. National Chinchilla Breeder 18, 20-22.
STURKIE P. D. (1965) Avian Physiology, p. 257. Cornell University Press, Ithaca, N.Y.
TURNER C. D. (1967) General Endocrinology, p. 5'79. W. B. Saunders, London.
Ключевые слова: шиншилла; самопогрызание меха; функция щитовидной железы; функция надпочечников; потребление кислорода; Chinchilla lanigera.
Авторы: W. J. VANJONACK and H. D. JOHNSON
Департамент молочного животноводства, Университет Миссури / Колумбия, Колумбия, миссури 65201
Получено 11 августа 1972 г.
Получено из Журнала сельскохозяйственных экспериментальных станций Миссури № 6384. Утверждено Директором.
Перевод: Д.В.Михайлов
Vanjonack W.J. and Johnson H.D. Relationship of thyroid and adrenal function to "fur-chewing" in the chinchilla (Взаимосвязь функции щитовидной железы и надпочечников с самопогрызанием меха у шиншилл) // Comp. Biochem. PhysioL, 1973, Vol. 45A, pp. 115 to 120. Pergamon Press. Printed in Great Britain
Файл с оригиналом статьи:
Все права защищены.
Использование материалов (текста и изображений) разрешается при условии ссылки на "Социальную сеть любителей шиншилл CHINBOX" (chinbox.online). Для интернет-изданий обязательной является прямая гиперссылка, не закрытая от индексации поисковыми системами (отсутствие в ссылке rel="nofollow" или ‹noindex› ), в первом или втором абзаце опубликованного материала (ссылка должна быть в тексте).