Аннотация
Самопогрызание меха является распространенной проблемой у шиншилл (Chinchilla lanigera). Это может затронуть благополучие животных по причине потери тепла, что может повлиять на потребление пищи и воды для поддержания температуры тела. В этом контексте инфракрасная термография представляется подходящим методом измерения тепловыделений от поверхности объектов.
Половозрелые домашние шиншиллы-самцы были разделены на две группы: «Без самопогрызания», демонстрирующие нормальное поведение (n = 25), и «с самопогрызанием», имевшие такую проблему (n = 23). Потребление пищи и воды (среднее значение ± стандартное отклонение), измеренное у животных контрольной группы, составило 20,7 ± 4,52 г и 15,9 ± 3,45 мл, в то время как у шиншилл с самопогрызанием оно было на 25% и 33% выше, соответственно. Теплопоток через погрызенные участки составлял 6,06 мВт / см2, что в 2,8 раза выше, чем через неповрежденный мех шиншилл. Подводя итог, тепловизионные изображения явно показывают, что самопогрызание меха вызывает повышенную потерю тепла. Нарушения поддержания теплового гомеостаза могут быть дополнительным фактором, снижающим благополучие этих животных.
Введение
Длиннохвостая шиншилла (Chinchilla lanigera, Molina, 1782) является популярным домашним и сельскохозяйственным животным. Естественная среда обитания диких шиншилл охватывает бесплодные, засушливые и холмистые районы поперечных горных цепей на севере центральной части Чили. Было предпринято несколько попыток одомашнить шиншиллу, но все нынешние колонии в неволе происходят от 12 диких особей, захваченных Мэтью Чепменом в 1920-х годах (Spotorno, Zuleta, Valladares, Deane, & Jimènez, 2004).
Самопогрызание меха (также известное как выкусывание меха) представляет собой одну из наиболее распространенных проблем при разведении шиншилл (Łapiński, Lis, Wójcik, Migdał, & Guja, 2014; Ponzio, Busso, Ruiz, & Fiol de Cuneo, 2007; Ponzio et al., 2012). Проявление такого поведения является одним из наиболее важных признаков плохого благосостояния в меховом хозяйстве (Broom & Fraser, 2007; WelFur, 2014). Его этиология не была окончательно установлена, но были рассмотрены различные причинные факторы, такие как условия проживания, психический стресс, основные заболевания и генетическая предрасположенность (Łapiński et al., 2014; Ponzio et al., 2007, 2012; Tisljar et al., 2002).
Одна из основных проблем в науках о животных связана с тем фактом, что инвазивные процедуры, такие как забор крови, вызывают стресс, снижают благосостояние животных и влияют на объективность результатов. Чтобы уменьшить эту проблему, был разработан ряд неинвазивных или минимально инвазивных методов и устройств (Choi, Miki, Sagawa & Shiraki, 1997; Peng et al., 2019; Stewart, Webster, Schaefer, Cook & Scott, 2005). Инфракрасная термография (IRT) представляет собой один из таких методов, который позволяет измерять теплоотдачу с поверхности объектов. Тепловыделение, которое отражается на температуре тела животных, связано со здоровьем и скоростью метаболизма (Choi et al., 1997; McCafferty, Moncrieff, Taylor, & Boddie, 1998; Montanholi et al., 2008; Peng et al. 2019). Физические теплоизоляционные свойства шерсти животных зависят от толщины, длины и плотности шерсти, поэтому нарушение структуры меха приводит к увеличению затрат энергии на поддержание надлежащей температуры тела (Forni et al., 2017; Hes, 2008). Представляется интересным определить, увеличивает ли самопогрызание меха потери тепла у шиншилл (Chinchilla lanigera).
Материалы и методы
Содержание и уход за животными
Половозрелые самцы (29–44 месяца) домашней шиншиллы (Chinchilla lanigera), стандартного окраса, были разделены на две группы: «без самопогрызания», демонстрирующие нормальное поведение (n = 25), и «самовыгрызающие», демонстрирующие самопогрызание меха (n = 23). Масса тела животных в двух группах (среднее значение ± стандартное отклонение) составила 550 ± 68,3 г и 594 ± 85,4 г соответственно. Особи с самопогрызанием меха были получены не менее чем за 6 месяцев до начала исследования из различных коммерческих племенных ферм и доставлены в питомник шиншилл Сельскохозяйственного университета в Кракове. Животные, имеющие нормальные поведенческие черты, происходили из того же учебного племенного хозяйства. Перед тестами обеим группам животных был предоставлен соответствующий период адаптации: они содержались в одном помещении в одинаковых условиях в течение 6 месяцев. Система содержания была такой же, как и на коммерческих шиншилловых фермах.
Животных содержали по отдельности в клетках из нержавеющей стали (ширина 0,40 м, длина 0,50 м, высота 0,35 м). Каждая клетка была оснащена полипропиленовыми поилками, самотёчным питателем из нержавеющей стали, купалкой (выдвижного типа) и деревянным буковым блоком для истирания зубов. Животных кормили 50 г / день гранулированной кормовой смесью «1790 Szynszyl-Raba» с метаболической энергией 9,6 МДж / кг (De Heus a. S, Таблица 1.) и давали 100 мл / день воды из полипропиленовой поилки. Во время исследования потребление пищи и воды контролировали каждые 24 часа путем взвешивания с точностью до 0,1 г. Суточное потребление корма определяли путем вычитания из начальной порции массы корма, оставшегося в кормушке и на поддоне под полом.
Животные подвергались воздействию контролируемой температуры (20–22 °C) и искусственного фотопериода (12 ч света / 12 ч темноты).
Параметры окружающей среды в виварии контролировали ежедневно в течение 11 дней исследования подряд: температуру и влажность измеряли с помощью аспирационного психрометра (термометр с влажным и сухим сосудом) и термоанемометра, тип TA465 (Test-Therm, Польша); скорость охлаждения измерялась с помощью сертифицированного кататермометра Hills (Kujawska Wytwórnia Termometrów, Польша; сертификат измерения: Институт метеорологии и управления водными ресурсами - Institute of Meteorology and Water Management – National Research Institute, Польша); концентрации CO2 и NH3 в воздухе измеряли с помощью многогазового детектора типа POLYTECTOR II G750 (GfG, Германия).
Таблица 1. Состав гранулированного корма для шиншилл «1790 Szynszyl-Raba» (De Heus A.S).
|
Компонент
|
Процентная доля, %
|
|
Сырой протеин
|
16,90
|
|
Сырое масло и жир
|
4,40
|
|
Сырая клетчатка
|
13,90
|
|
Сырая зола
|
7,50
|
|
Лизин
|
0,84
|
|
Метионин
|
0,37
|
|
Кальций
|
0,80
|
|
Фосфор
|
0,62
|
|
Натрий
|
0,25
|
Все животные в исследовании привыкли к уходу и одинаковому обращению, чтобы унифицировать любые индивидуальные различия; эксперименты проводились в соответствии с одобрения Первого местного этического комитета по испытаниям на животных при Ягеллонском университете в Кракове (45/2014). Инфракрасную термографию проводили с использованием камеры A20 с разрешением детектора 120 × 160 пикселей и излучательной способностью ɛ = 0,98 (FLIR Systems Inc., США). Следует отметить, что инфракрасные измерения характеризуются очень точным определением разницы температур между двумя точками (ΔT), в то время как измерение значения температуры не всегда является верным (Więcek & De Mey, 2011). Температура окружающей среды (минимальная температура окружающей среды, ta), температура поверхности тела в выбранной области (ts), температура контрольных точек (т. е. самая высокая зарегистрированная температура в области глаза или ануса, tr) и перепады температур ( Δtsa, Δtra и Δtrs) определяли с использованием программного обеспечения FLIR ThermaCAM Researcher. Камера была установлена на штатив на расстоянии 75 см от животного, таким образом поле обзора камеры под углом 45 градусов включало сечение 40 см, это гарантировало, что на долю шиншиллы, помещенной в центре камеры, приходится менее 30 градусов.
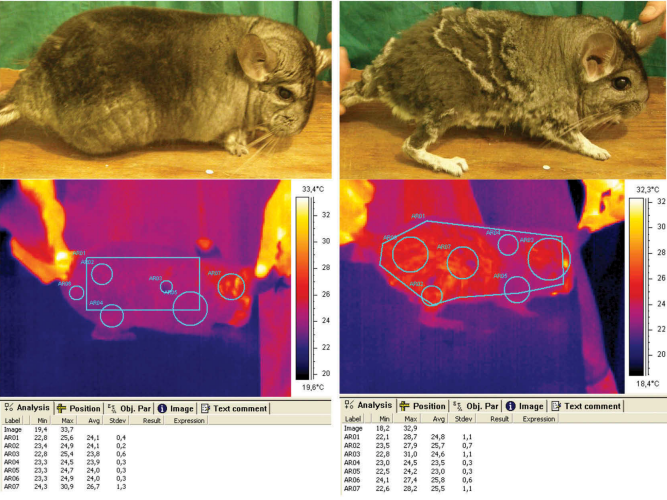
Термографические измерения проводились в комнате, где содержались животные. Для каждого животного была сделана серия из 6 термограмм, включавших следующие части тела: левую и правую стороны, спину, живот, переднюю и заднюю части животного. Продолжительность серии составила около 1 мин. Каждую термограмму в серии анализировали индивидуально, а затем параметры усредняли для каждого животного. Шиншиллы были расположены таким образом, чтобы плоскость измерения была полностью видна на термограмме (рис. 1). Все поверхности на левой и правой стороне тела, спине и животе каждой шиншиллы с видимым повреждением меха были классифицированы как выгрызенные. Фотографии особей сравнивались с одновременно снятыми термограммами (рис. 1). На основе этого была определена зона самопогрызания. Процент выгрызенной поверхности на изображениях был рассчитан как отношение количества пикселей в погрызенной области к количеству пикселей в общей площади в соответствии с формулой. (1):
где:
RC - относительная площадь самопогрызания [%];
S ch - область самопогрызания меха на термограмме животного [пиксель];
S t - общая площадь термограммы животного [пиксель];
Количество пикселей было определено с помощью Gimp 2.8.14.1 (GNU Image Manipulation Program)
Рисунок 1. Сравнение фотографий и термограмм шиншиллы из контрольной группы (слева) и шиншиллы с самопогрызанием меха (справа).
Расчет теплопотери
Данные, полученные термографически, были использованы для расчета потерь тепла (Q [W]) через поверхность с нормальной шерстью и с самопогрызанием, с использованием формулы, предложенной McCafferty et al. (2011) (2):
где:
A - средняя площадь поверхности тела шиншиллы, A = 0,0496 м2 (Barabasz, Łapiński, & Grembowicz, 2010);
t s - температура поверхности меха [° C] по данным термограммы;
t a - температура воздуха [° C] по данным термограммы;
h - коэффициент теплопередачи [Вт м-2 ° С-1].
Коэффициент теплопередачи кожи шиншиллы определяли в лаборатории (t a = 23ºC) с использованием кататермометра Hills, обернутого в дубленую кожу шиншиллы с нормальным или выгрызенным мехом, и рассчитывали по следующей формуле (3):
Где:
q - индекс ката [Вт м-2]; то есть потери тепла через поверхность колбы, рассчитанные путем измерения времени охлаждения [s] кататермометра, сертифицированного с индивидуальным коэффициентом [Дж дм-2] от 38 до 35ºC (Parson, 2014);
Δt r-a - разница температур между контрольными точками (глаз и анус) и температурой окружающей среды, определенная по термограммам (составила 13,8 ± 1,18ºC).
Статистический анализ
Влияние самопогрызания меха на потерю тепла и потребление пищи и воды было проверено с помощью одностороннего ANOVA. Пост-хок анализ проводился с использованием критерия Тьюки. Также была смоделирована Взаимосвязь между RC [%] и Q [W]. Статистический анализ проводился с использованием Sigma-Stat 3.5 (SPSS Science Software Ltd., США), а диаграммы подготовлены с помощью Grapher 8.0 (Golden Software Inc., США).
Результаты
Условия окружающей среды
Температурно-влажностные условия во время эксперимента были стабильными: температура (среднее значение ± стандартное отклонение) 20,6 ± 0,62ºC; относительная влажность 50 ± 7,7%; скорость охлаждения 196 ± 9,4 Вт м -2 °С -1; концентрация СО2 в воздухе 0,03%; Уровень NH3 был ниже чувствительности прибора.
Питание и потребление воды
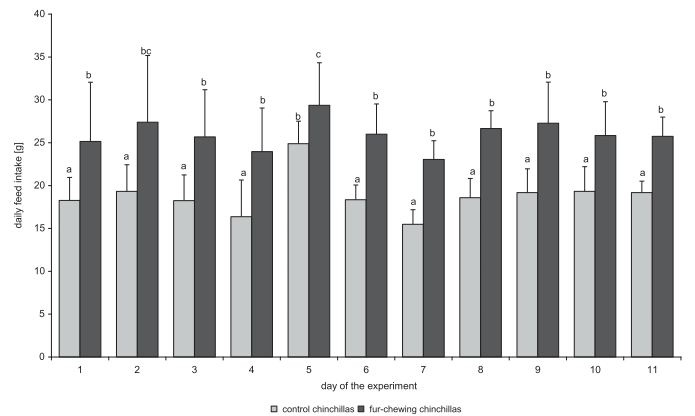
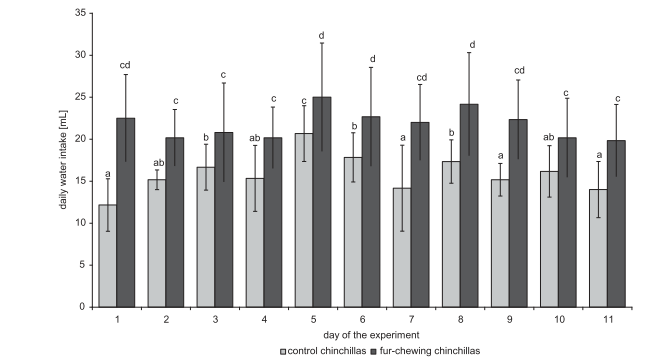
Потребление пищи и воды (среднее значение ± стандартное отклонение) у животных контрольной группы составило 20,7 ± 4,52 г и 15,9 ± 3,45 мл соответственно, в то время как у шиншилл с самопогрызанием меха эти значения составили 25,8 ± 3,20 г и 21,1 ± 4,83 мл соответственно (P < 0,05; рисунки 2 и 3). Такое же соотношение сохранялось для относительного потребления пищи (контрольная группа, 37,4 ± 5,87 г / кг массы тела; шиншиллы с самопогрызанием меха, 43,8 ± 6,90 г / кг массы тела) и воды (контрольная группа, 29,3 ± 7,76 мл / кг массы тела; шиншиллы с самопогрызанием меха, 35,9 ± 7,53 г / кг массы тела) (р <0,05). Потребление энергии у контрольной и группы шиншилл с самопогрызанием, рассчитанное с использованием приведенных выше данных и информации по кормовой энергии, составило 2,2 и 2,8 Вт соответственно.
Рисунок 2. Ежедневное потребление пищи (среднее значение ± стандартное отклонение) контрольной группы и группы шиншилл с самопогрызанием.
Прим. пер. к рис. 2: daily feed intake [mL] – потребление корма в день (мл), day of the experiment – день эксперимента, control chinchillas – шиншиллы контрольной группы, fur-chewing chinchillas – шиншиллы с самопогрызанием меха.
Рисунок 3. Ежедневное потребление воды (среднее значение ± стандартное отклонение) контрольной группы и группы шиншилл с самопогрызанием.
Прим. пер. к рис. 2: daily water intake [mL] – потребление воды в день (мл), day of the experiment – день эксперимента, control chinchillas – шиншиллы контрольной группы, fur-chewing chinchillas – шиншиллы с самопогрызанием меха.
Оценка кожного теплообмена
Индекс ката колбы, измеренный в лаборатории (среднее значение ± стандартное отклонение), составлял 173,3 ± 10,58 Вт м-2, в то время как индекс ката через кожу шиншиллы составлял 69,8 ± 8,43 Вт м-2, а через снятую кожу шиншиллы - 129,9 ± 13,53 Вт м -2. Исходя из этого, были определены коэффициенты теплопередачи для кожи шиншилл контрольной группы и кожи шиншилл с самопогрызанием меха: 5,06 ± 0,611 Вт м -2 °С-1 (0,506 мВт см-2 °С-1) и 9,41 ± 0,902 Вт м-2 °С-1 (0,941 мВт / см 2 °C) [таблица 2].
Анализ термограмм показал, что животные в нашем эксперименте имели различные уровни интенсивности самопогрызания меха. Площадь поверхности кожи с выгрызенным мехом (среднее значение ± стандартное отклонение) составляла 12,8 ± 16,44% и варьировалась от 0,8% (незначительные, только несколько выгрызенных волос) до 43,2% (очень тяжелая степень, выгрызен весь мех в областях, которые могут быть достигнуты животным).
Анализ термограмм показал, что и глаз, и анус выделялись на изображениях как «горячие точки». Различия между температурой поверхности кожи и глаз или ануса были одинаковыми (8,0 ± 1,58°C и 7,6 ± 1,06°C соответственно); следовательно, они могут быть использованы в качестве ориентиров. Кроме того, не было обнаружено различий в Δtr-a между контрольной группой (13,8 ± 1,02°C) и группой шиншилл с самопогрызанием (13,8 ± 1,21°C).
Тепловой поток через погрызенные участки был рассчитан как 6,06 ± 0,796 мВт / см2, что в 2,8 раза выше, чем через неповрежденный мех шиншилл (2,19 ± 0,296 мВт / см2, P <0,05, таблица 2). Общие потери тепла со всей поверхности тела у животных, выгрызавших мех, были на 21% выше, чем у контрольных животных (P <0,05, таблица 2). Зависимость между областями с выгрызенным мехом (x), потреблением пищи (y) и потреблением воды (z) может быть описана следующими уравнениями: y = 0,245x + 21,646 (R2 = 0,28); z = 0,244х + 16,939 (R2 = 0,31).
Обсуждение
Благополучие животных можно оценить по четырем принципам: хорошее жилье, хорошее питание, хорошее здоровье и соответствующее поведение. Параметры окружающей среды на объекте размещения шиншилл были выбраны на основе сформулированных рекомендаций (T = 14–30°C, RH = 45–55%) Tombarkiewicz, Szeleszczuk, Niedziółka, and Rapacz (2003). Температурный диапазон (20,6 ± 0,62°C), использованный во время эксперимента, находился в пределах термонейтральной зоны для взрослой шиншиллы, оцененной Drożdż и Gorecki (1967), которая колеблется от 20ºC до 30ºC, но был несколько ниже, чем диапазон (между 22ºC и 35ºC), установленный Arends and McNab (2001). Ухудшение термического гомеостаза вызывает изменения метаболизма, которые проявляются в виде изменений потребления пищи и воды (Schmidt-Nielsen, 1997). Однако во время исследования потребление пищи и воды в обеих группах находилось в нижнем диапазоне нормы (около 55 г корма на кг массы тела и 20–120 мл воды на кг массы тела), рекомендованной Fehr (2008) и Hagen, Clauss, и Hatt (2014), а потребление воды, по-видимому, зависело от потребления корма (в обеих группах одинаково, т. е. 0,80–0,82 мл воды на 1 г корма). Различия в потреблении энергии могут также зависеть от других поведенческих расстройств. Объективно следует отметить, что происхождение животных из разных популяций и их предыстория и среда обитания (переселение в университетский питомник из различных коммерческих ферм) могут влиять как на общие уровни активности, так и на самопогрызание меха. Известно, что у большинства видов грызунов наблюдается аномальное повторяющееся поведение, когда они содержатся в скудных по наполнению клетках (Würbel, 2006). Индивидуальные различия означают, что у некоторых животных развивается аномальное повторяющееся поведение, в то время как другие становятся пассивными, что может отражать активный и пассивный тип личности. Franchi, Alejandro и Tadich (2016) установили, что эти расстройства поведения также относятся и к шиншиллам. С другой стороны, в том же исследовании не было обнаружено существенных различий между временным бюджетом или количеством событий в день для каждого зарегистрированного аномального повторяющегося поведения контрольных и самопогрызающих мех шиншилл.
На теплообмен через кожу влияют два типа факторов: физические и физиологические. Физиологические факторы сильно различаются, поскольку кровообращение, особенно вазоконстрикция капилляров в сравнении с вазодилатацией, когда есть необходимость сберегать либо рассеивать тепло, составляет существенную часть механизмов теплопередачи живой кожи. Физиологическим ответом, вызываемым холодом, является вазоконстрикция и ограниченный кровоток (Schmidt-Nielsen, 1997). Аналогичная реакция была описана Forni et al. (2017) у частично обритых мышей. Однако у животных, подвергающихся дополнительному стрессу (ограничение калорийности), базальный кровоток был увеличен, что указывает на то, что кожная сосудистая система в этой группе имеет более низкую степень вазоконстрикции по сравнению с ожидаемой. Это может дополнительно объяснить повышенную потерю тепла, наблюдаемую у ограниченных в калориях животных по сравнению с животными, которых кормили ad libitum.
Таблица 2. Потери тепла (Q) для шиншилл с самопогрызанием меха, рассчитанные по термограммам. Разница (ts-a) между температурой поверхности меха (ts) и температурой окружающей среды (ta) были рассчитаны по термограммам. Коэффициент теплопередачи меха (h) был определен при лабораторных измерениях кожи шиншиллы. Площадь кожи шиншиллы (A,) определялась на основе протокола, разработанного Barabasz et al. (2010), а площадь самопогрызания меха была рассчитана с использованием термограмм. Значения были представлены как среднее ± стандартное отклонение (SD).
|
Животные
|
Анализируемая поверхность
|
ts [ºC]
|
ta [ºC]
|
Δts-a [ºC]
|
h[mW см−2°C−1]
|
Тепловой поток h× Δts-a [мВт см− 2]
|
A [cm2]
|
Q [мВт]
|
|
Контрольная группа
|
Мех целиком (n=23)
|
23.7 ± 0.28 a
|
18.9 ± 0.44
|
4.9 ± 0.43 a
|
0.506
|
2.28 ± 0.178a
|
496 ± 52.9
|
1132 ± 88.3 a
|
|
Группа шиншилл с самопогрызанием
|
Мех целиком (n=25)
|
24.3 ± 0.59 b
|
18.9 ± 0.38
|
5.5 ± 0.70 b
|
0.506
|
2.76 ± 0.352b
|
496 ± 52.9
|
1372 ± 174.9 b
|
|
Невыгрызенные области (n=6)
|
23.3 ± 0.32 a
|
19.0 ± 0.65
|
4.3 ± 0.59 a
|
0.506
|
2.19 ± 0.296 a
|
432 ± 81.5
|
953 ± 244.0*
|
|
Выгрызенные области (n=6)
|
25.4 ± 1.06 b
|
19.0 ± 0.71
|
6.4 ± 0.85 b
|
0.941
|
6.06 ± 0.796 b
|
64 ± 81.5
|
386 ± 292.6*
|
Значения ab в столбцах, отмеченных различными буквами, значимо различаются (P <0,05).
* - значения не сопоставимы из-за того, что относятся к разным поверхностям
Для упрощения модели в настоящем исследовании, было сделано предположение, что живая кожа обладает свойствами теплообмена, подобными дубленой коже мертвого животного. Эти предположения основаны на следующих факторах: 1) дерма шиншиллы очень тонкая (толщиной около 400 мкм), а ее меховой покров является основным фактором сохранения тепла (Ptak, 1975; Wilcox, 1950), p. 2) C. lanigera демонстрирует самое высокое значение теплоизоляции, зарегистрированное у грызунов, и потери воды при испарении у этих животных очень низки - 95% от ожидаемого значения для мешотчатопрыгуновых (Cortes, Rosenmann, & Bozinovic, 2000). Поэтому было принято, что физические факторы были более важны, чем физиологические факторы, которые по этой причине были исключены.
В нашем исследовании средняя площадь самопогрызания меха составила 12,8% поверхности тела, что привело к приблизительному увеличению теплопередачи на 21% и увеличению потребления энергии с кормом на 27%. Эти результаты соответствуют работе Forni et al. (2017), в которой было подсчитано, что частичное бритье меха привело к увеличению расхода энергии у 24% мышей, которых кормили ad libitum, и у 40% с ограничением по калориям. Участки с погрызенным мехом отличаются короткими и редкими волосами, или вообще не имеют шерсти в тяжелых случаях. Чтобы поддерживать стабильную температуру, тело должно компенсировать потерю тепла путем увеличения выработки метаболической энергии (Schmidt-Nielsen, 1997).
Потребление кислорода у шиншилл при температуре 20 °C составляет 0,6–0,7 см3 г -1 ч -1, а выработка тепла - до 32 ккал / животное / сутки (Arends & McNab, 2001; Drożdż & Gorecki, 1967), что эквивалентно ок. 1550–1650 мВт. Эти авторы также утверждают, что меховой покров в отличном состоянии снижает потери тепла на 6–10%. Данные теплового потока, полученные из анализа термограмм, соответствуют приведенным выше наблюдениям.
Анализ тепловыделения был выполнен на основе метода, предложенного McCafferty et al. (2011), который позволяет рассчитывать тепловыделение, используя только информацию о температуре поверхности, когда невозможно измерить внутреннюю температуру тела (объекта). Температура поверхности тела у шиншилл определялась на основе средних температур, измеренных в областях, обозначенных с одной стороны животного. Высокая плотность шерсти и малый размер тела этих грызунов приводят к выравниванию температуры поверхности грудной клетки шиншилл контрольной группы. На этом фоне выделяются участки, не покрытые мехом (то есть глаза, уши и анус) (Polit, Rząsa, Rafajłowicz & Ni ,ański, 2018). Эти области с температурой, подобной внутренней температуре тела, были предложены некоторыми авторами в качестве ориентиров для использования при анализе термограмм (Łapiński et al. 2011, Nääs et al., 2014, Polit et al. 2018).
Самопогрызание меха, несомненно, является симптомом снижения благосостояния (Broom & Fraser, 2007; Franchi et al., 2016; Ponzio et al., 2007, 2012; WelFur, 2014). Кроме того, это способствует увеличению тепловыделения от поверхности тела. Теплоизоляция, обеспечиваемая мехом, зависит от длины и густоты меха (Morgan, 1997); поэтому для каждой породы животных характерна определенная термически нейтральная зона (Autio, Neste, Airaksinen & Heiskanen, 2006). Интенсивный теплообмен создает ощущение холода, которое может вызывать дискомфорт, влиять на поведение и снижать благосостояние животных (Karp, 2012). Увеличенное потребление корма компенсирует потребность в метаболической энергии, но не обязательно снижает психологический эффект.
Заключение
Термографические измерения ясно показывают увеличение теплоотдачи через кожу, лишенную изолирующего слоя в результате самопогрызания меха; это может отрицательно сказаться на тепловом комфорте животных и еще больше снизить их благосостояние. Из-за увеличенного потребления корма шиншиллам с самопогрызанием меха должно быть предоставлено достаточно пищи для поддержания температуры тела. Инфракрасная технология и тепловидение стали популярными инструментами для исследования терморегуляции у животных. Инфракрасная термография помогает нам помочь понять то, как животные приспосабливаются к окружающей среде, поскольку позволяет изучать механизмы рассеивания и сохранения тепла. Более того, дальнейшее развитие этого метода исследования и сочетание с метаболическим анализом может предоставить чувствительный инструмент для оценки параметров благополучия животных.
Соответствие этическим нормам
Конфликт интересов. Авторы заявляют, что не имеют конфликта интересов.
Одобрение этического комитета
Для данного типа исследования формальное согласие не требуется.
Финансирование
Это исследование финансировалось Министерством науки и высшего образования (DS-3263 и DS 3266).
Использованные источники
Arends, A., & McNab, B. K. (2001). The comparative energetics of ‘caviomorph’ rodents. Comparative Biochemistry and Physiology, Part A, 130, 105–122.
Autio, E., Neste, R., Airaksinen, S., & Heiskanen, M. L. (2006). Measuring the heat loss in horses in different seasons by infrared thermography. Journal of Applied Animal Welfare Science, 9, 211–221.
Barabasz, B., Łapiński, S., & Grembowicz, M. (2010). Zależności pomiędzy parametrami fenotypowymi, masą ciała i wielkością skóry szynszyli [The relationship between phenotypic parameters, body weight and skin size of chinchillas]. Roczniki Naukowe Polskiego Towarzystwa Zootechnicznego, 6, 113–119. in Polish.
Broom, D. M., & Fraser, A. F. (2007). Domestic animal behaviour and welfare (4th ed.). Wallingford, Oxfordshire, UK: CABI.
Choi, J. K., Miki, K., Sagawa, S., & Shiraki, K. (1997). Evaluation of mean skin temperature formulas by infrared thermography. International Journal of Biometeorology, 41, 68–75.
Cortes, A., Rosenmann, M., & Bozinovic, F. (2000). Relación costo_beneficio en la termorregulación de Chinchilla lanigera [Cost-benefit relationship in thermoregulation of Chinchilla lanigera]. Revista Chilena De Historia Natural, 73, 351–357.
Drożdż, D., & Gorecki, A. (1967). Oxygen consumption and heat production in chinchillas. Acta Theriologica, 12, 81–86.
Fehr., M. (2008). Chinchilla. In K. Gabrisch & P. Zwart (Eds.), Krankheiten der heimtiere (pp. 183–212). Hannover: Schlutersche.
Forni, M. F., Peloggia, J., Braga, T. T., Chinchilla, J. E. O., Shinohara, J., Navas, C. A., … Kowaltowski, A. J. (2017). Caloric restriction promotes structural and metabolic changes in the skin. Cell Reports, 20, 2678–2692.
Franchi, V., Alejandro, A. O., & Tadich, T. A. (2016). Fur chewing and other abnormal repetitive behaviors in chinchillas (Chinchilla lanigera), under commercial fur-farming conditions. Journal of Veterinary Behavior, 11, 60–64.
Hagen, K., Clauss, M., & Hatt, J. M. (2014). Drinking preferences in chinchillas (Chinchilla laniger), degus (Octodon degu) and guinea pigs (Cavia porcellus). Journal of Animal Physiology and Animal Nutrition, 98, 942–947.
Hes, L. (2008). Analysing the thermal properties of animal furs for the production of artificial furs. In I. A. Abbott & M. Ellison (Eds.), Biologically inspired textiles (pp. 150–167). Cambridge: Woodhead Publishing.
Karp, C. L. (2012). Unstressing intemperate models: How cold stress undermines mouse modeling. Journal of Experimental Medicine, 209, 1069–1074.
Łapiński, S., Augustyn, J., Lis, M. W., Kanik, W., Niedbała, P., & Lisowska-Lis, A. (2011). Zmiany w obrazie termo- graficznym królika domowego (Oryctolagus cuniculus) w zależności od wieku i stanu fizjologicznego [Changes in domestic rabbit (Oryctolagus cuniculus) thermographic image, depending on its age and physiological condition]. Pomiary, Automatyka, Kontrola, 57, 1154–1157.
Łapiński, S., Lis, M. W., Wójcik, A., Migdał, Ł., & Guja, I. (2014). Analysis of factors increasing the probability of fur chewing in chinchilla (Chinchilla laniegera) raised under farm conditions. Annals of Animal Science, 14, 189–195.
McCafferty, D. J., Gilbert, C., Paterson, W., Pomeroy, P. P., Thompson, D., Currie, J. I., & Ancel, A. (2011). Estimating metabolic heat loss in birds and mammals by combining infrared thermography with biophysical modeling. Comparative Biochemistry and Physiology, Part A, 158, 337–345.
McCafferty, D. J., Moncrieff, J. B., Taylor, I. R., & Boddie, G. F. (1998). The use of IR thermography to measure the radiative temperature and heat loss of a barn owl (Tyto alba). Journal of Thermal Biology, 23, 311–318.
Montanholi, Y. R., Odongo, N. E., Swanson, K. C., Schenkel, F. S., McBride, B. W., & Miller, S. P. (2008). Application of infrared thermography as an indicator of heat and methane production and its use in the study of skin temperature in response to physiological events in dairy cattle (Bos taurus). Journal of Thermal Biology, 33, 468–475.
Morgan, K. (1997). Thermal insulance of peripheral tissue and coat in sport horses. Journal of Thermal Biology, 22, 169–175. Nääs, I. A., Garcia, R. G., & Caldara, F. R. (2014). Infrared thermal image for assessing animal health and welfare. Journal Of Animal Behaviour and Biometeorology, 2, 66–72.
Parson, K. (2014). Measurement methods and assessment techniques. Human thermal environments, the effects of hot: Moderate, and cold environments on human health, comfort, and performance (3rd ed., pp. 101–138).Boca Raton: CRC Press & Taylor & Francis Group..
Peng, D., Chen, S., Li, G., Chen, J., Wang, J., & Gu, X. (2019). Infrared thermography measured body surface temperature and its relationship with rectal temperature in dairy cows under different temperature-humidity indexes. International Journal of Biometeorology. doi:10.1007/s00484-018-01666-x
Polit, M., Rząsa, A., Rafajłowicz, W., & Niżański, W. (2018). Infrared technology for estrous detection in Chinchilla lanigera. Animal Reproduction Science, 197, 81–86.
Ponzio, M. F., Busso, J. M., Ruiz, R. D., & Fiol de Cuneo, M. (2007). A survey assessment of the incidence of fur chewing in commercial chinchilla (Chinchilla lanigera) farms. Animal Welfare, 16, 471–479.
Ponzio, M. F., Monfort, S. L., Busso, J. M., Carlini, V. P., Ruiz, R. D., & Fiol de Cuneo, M. (2012). Adrenal activity and anxiety-like behaviour in fur-chewing chinchillas (Chinchilla lanigera). Hormones and Behavior, 61, 758–762.
Ptak, W. (1975). Budowa histologiczna skory szynszyli (Chinchilla velligera Prell 1934) i budowa naskorka i stosunek jego grubosci do grubosci skory właściwej [Structure of the epidermis and its relation to the thickness of the derma]. Zeszyty Naukowe AR Kraków, 98, Zootechnika, 15, 49–74.
Schmidt-Nielsen, K. (1997). Animal physiology, adaptation and environment (Fifth). (pp. 302–367). Cambridge University Press. (Polish ed. 2008. . Warszawa: PWN.
Spotorno, E., Zuleta, C. A., Valladares, J. P., Deane, A. L., & Jimènez, J. E. (2004). Chinchilla Laniger. Mammalian Species, 758, 1–9.
Stewart, M., Webster, J. R., Schaefer, A. L., Cook, N. J., & Scott, S. L. (2005). Infrared thermography as a non-invasive tool to study animal welfare. Animal Welfare, 14, 319–325.
Tisljar, M., Janić, D., Grabarević, Z., Simpraga, B., Marinculić, A., Pinter, L., … Nemanic, A. (2002). Stress-induced Cushing’s syndrome in fur-chewing chinchillas. Acta Veterinaria Hungarica, 50, 133–142.
Tombarkiewicz, B., Szeleszczuk, O., Niedziółka, J., & Rapacz, A. (2003). Warunki zoohigieniczne pomieszczenia a wskaźniki użytkowości rozpłodowej szynszyli (Chinchilla velligera) [Hygienic conditions in cages and reproductive performance of the chinchilla (Chinchilla velligera)]. Prace Komisji Nauk Rolniczych I Biologicznych. BTN B, 51, 275–280.
WelFur. (2014). WelFur. Welfare assesmant protocol for foxes (pp. 50–51). Brussels: Author.
Więcek, B., & De Mey, G. (2011). Termowizja w podczerwieni, podstawy i zastosowania [Infrared thermovision, foundations and applications]. Warsaw: Wyd. PAK.
Wilcox, H. H. (1950). Histology of the skin and hair of the adult chinchilla. The Anatomical Record, 108, 385–397.
Würbel, H. (2006). The motivational basis of caged rodents’ stereotypies. In G. Mason & J. Rushen (Eds.), Stereotypic animal behaviour, fundamentals and applications to welfare (2nd ed., pp. 86–120). London UK: CABI.
Авторы: Stanisław Łapiński a, Joanna Orel b, Piotr Niedbała a, Weronika Kucharskab, Magdalena Jakubowska b, Agnieszka Lisowska-Lis c, Barbara Tombarkiewiczb , and Marcin W. Lis b
a Department of Swine and Small Animal Breeding, University of Agriculture in Kraków, Kraków, Poland; b Departmentof Veterinary, Reproduction and Animal Welfare, Faculty of Animal Science, University of Agriculture in Kraków,Kraków, Poland; c State Higher Vocational School in Tarnów, Polytechnic Institute, Tarnów, Poland.
JOURNAL OF APPLIED ANIMAL WELFARE SCIENCE, https://doi.org/10.1080/10888705.2019.1614924
Перевод на русский язык: Д.В. Михайлов
Stanisław Łapiński, Joanna Orel, Piotr Niedbała, Weronika Kucharska, Magdalena Jakubowska, Agnieszka Lisowska-Lis, Barbara Tombarkiewicz, and Marcin W. Lis. Infrared Thermography as an Indicator of Heat Loss in Fur-Chewing Chinchillas (Chinchilla Lanigera) // J Appl Anim Welf Sci. 2019 May 16:1-10.